Anatomy and Physiology
Copyright ©
March 2003 Ted Nissen
Table
of Contents
8 Skeletal System-Appendicular Skeleton*
13 Spinal Cord and Spinal Nerves*
14 The Brain and Cranial Nerves
15 Sensory, Motor, and Integrative
Systems
19 Cardiovascular System-Blood*
20 Cardiovascular System-Heart
21 Cardiovascular System-Vessels and Routes
22 Lymphatic and Immune System*
27 Fluid, Electrolyte, and
Acid-Base Dynamics
29 Development and Inheritance
1 Introduction
2 Chemical Organization
2.1 General Introduction
2.1.1 Many of the common substances we eat and drink-water, sugar, table salt, cooking oil-play vital roles in keeping us alive. In this chapter, you will learn something about how these substances function in your body. Fundamental to this study is knowledge of basic chemistry and chemical processes, since your body is composed of chemicals and all body activities are chemical in nature. To understand the nature of the matter you are made from and the changes this matter goes through in your body, you will need to know which chemical elements are present in the human organism and how they interact.
2.2 Introduction to Basic Chemistry
2.2.1 Chemical Elements
2.2.1.1 All living and nonliving things consist of matter, which is anything that occupies space and has mass. Matter may exist in a solid, liquid, or gaseous state. All forms of matter are made up of a limited number of building units called chemical elements, substances that cannot be decomposed into simpler substances by ordinary chemical reactions. At present, scientists recognize 106 different elements, of which 92 occur naturally. Elements are designated by letter abbreviations, usually derived from the first or first and second letters of the Latin or English name for the element. Such letter abbreviations are called chemical symbols. Examples of chemical symbols are H (hydrogen), C (carbon), 0 (oxygen), N (nitrogen), Na (sodium), K (potassium), Fe (iron), and Ca (calcium).
2.2.1.2 Periodic Table Web Reference
2.2.1.2.1 http://www.webelements.com/index.html
2.2.1.2.2 http://www.periodic-table.com/
2.2.1.3 Approximately 26 elements are found in the human organism. Oxygen, carbon, hydrogen, and nitrogen make up about 96 percent of the body's weight. These four elements together with calcium and phosphorous constitute approximately 99 percent of the total body weight. Twenty other chemical elements, called trace elements, are found in low concentrations and compose the remaining 1 percent.
2.2.2 Particle Physics Link
2.2.2.1 Particle Physics
2.2.3 Structure of Atoms
2.2.3.1 Each element is made up of units of matter called atoms, the smallest units of matter that enter into chemical reactions. An element is simply a quantity of matter composed of atoms all of the same type. A handful of the element carbon, such as pure coal, contains only carbon atoms. A tank of oxygen contains only oxygen atoms. Measurements indicate that the smallest atoms are less than 0.00000001 cm 1/250,000,000 inch) in diameter, and the largest atoms are 0.00000005 cm (1/50,000,000 inch) in diameter. In other words, if 50 million of the largest atoms were placed end to end, they would measure approximately 2.5 cm (1 inch) in length.
2.2.3.2 An atom consists of two basic parts: the nucleus and electrons (Figure 2-1).
2.2.3.3 Illustration (Figure 2-1)
2.2.3.3.1 Introduction
2.2.3.3.1.1 Structure of an atom. In this highly simplified version of a carbon atom, note the centrally located nucleus. The nucleus contains six neutrons and six protons, although all are not visible in this view since some are behind others. The six electrons move about the nucleus at varying distances from its center
2.2.3.3.1.2 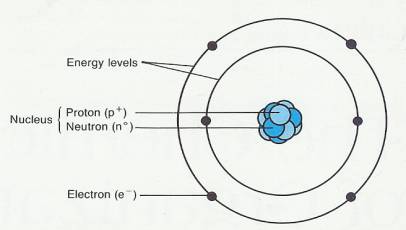
2.2.3.4 The centrally located nucleus constitutes most of the atomic mass and contains positively charged particles called protons (p+) and uncharged (neutral) particles called neutrons (nO). Because each proton has one positive charge, the nucleus itself is positively charged. Together, protons and neutrons are referred to as particles called nucleons. Various coordinated movements of nuclear particles cause the nucleus to vibrate in several distinctive patterns. Electrons (e-) are negatively charged particles that move around the nucleus. For reasons far beyond the scope of this text-. book, the diagrams of atoms are very simplified, resembling planetary models of the solar system. (In actuality, electrons do not follow fixed paths around the nucleus but move about in probable locations.) The number of electrons in an atom of an element always equals the number of protons. Since each electron carries one negative charge, the negatively charged electrons and the positively charged protons balance each other, and the atom is electrically neutral.
2.2.3.5 The standard relative weight unit for measuring subatomic particles and atoms is called an atomic mass unit (amu) or dalton. A neutron has a mass of 1.008 daltons, and a proton has a mass of 1.007 daltons. The mass of an electron is 0.0005 daltons, about 1/2000 the mass of a neutron or proton.
2.2.3.6 What makes the atoms of one element different from those of another? The answer lies in the number of protons. Figure 2-2 shows that the hydrogen atom contains one proton; the helium atom contains two; the carbon atom has six; and so on. Each different kind of atom has a different number of protons in its nucleus. The number of protons in an atom is called the atom's atomic number. Therefore, we can say that each kind of atom, or element, has a different atomic number. The total number of protons and neutrons in an atom is its approximate atomic weight. Each proton and neutron contributes one dalton unit of weight to the atom. Thus, an atom of sodium has an atomic weight of23 because of the presence of 11 protons and 12 neutrons in its nucleus.
2.2.3.7 Illustration (Figure 2-2) Atomic Structures of some Representative Atoms
2.2.3.7.1 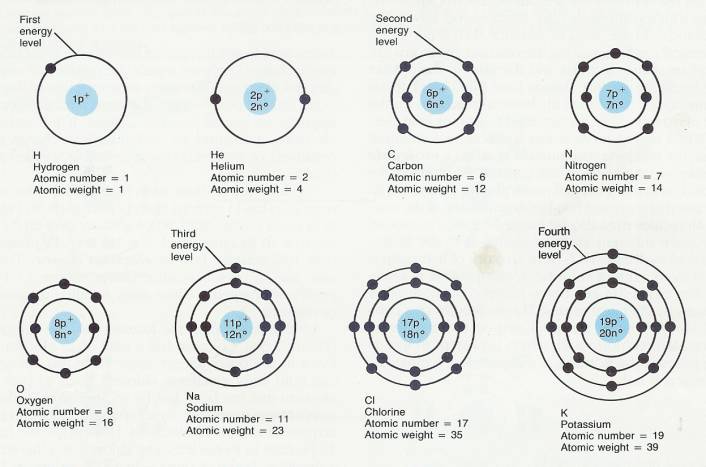
2.2.3.8 Atoms of an element, although chemically alike, may have different nuclear masses and thus different atomic weights because of one or more extra neutrons in some of the atoms, so the atomic weight assigned to an element is only an average. Each of the chemically identical atoms of an element with a particular nuclear mass is an isotope of that element. All isotopes of an element have the same number of protons in their nuclei, but their atomic weights differ because of the difference in the number of neutrons. In a sample of oxygen, for example, most atoms have 8 neutrons, but a few have 9 or 10, even though all have 8 protons. The isotopes of oxygen are designated as 160, 170, and 180. The numbers indicate their atomic weights.
2.2.3.9 Certain isotopes called radioisotopes are unstable they "decay" or change their nuclear structure to a more stable configuration. And in decaying they emit high energy radiation (alpha, beta, or gamma particles) that can be detected by instruments. These instruments estimate the amount of radioisotope present in a part of the body or in a sample of material and form an image of its distribution. (Refer to the discussion of positron emission tomography, or PET, in Chapter 1.)
2.2.4 Atoms and Molecules
2.2.4.1 When atoms combine with or break apart from other atoms, a chemical reaction occurs. In the process, new products with different properties are formed. Chemical reactions are the foundation of all life processes. .
2.2.4.2 The electrons of an atom actively participate in chemical reactions. The electrons move around the nucleus in regions, shown in Figure 2-2 as concentric circles lying at varying distances from the nucleus. We call these regions energy levels. Each energy level has a maximum number of electrons it can hold. For instance, the energy level nearest the nucleus never holds more than two electrons, no matter what the element. This energy level can be referred to as the first energy level. The second energy level holds a maximum of eight electrons. The third level of atoms whose atomic number. is less than 20 also can hold a maximum of eight electrons. The third level of more complex atoms can hold a maximum of 18 electrons.
2.2.4.3 An atom always attempts to fill its outermost energy level with the maximum number of electrons it can hold. To do this, the atom may give up, take on, or share electrons with another atom-whichever is easiest. The valence (combining capacity) is the number of extra or deficient electrons in the outermost energy level. Take a look at the chlorine atom. Its outermost energy level, which happens to be the third level, has seven electrons. Since the third level of an atom can hold a maximum of eight electrons, chlorine can be described as having a shortage of one electron. In fact, chlorine usually does try to pick up an extra electron. Sodium, by contrast, has only one electron in its outer level. This again happens to be the third energy level. It is much easier for sodium to get rid of the one electron than to fill the third level by taking on seven more electrons. Atoms of a few elements, like helium, have completely filled outer energy levels and do not need to gain or lose electrons. These are called inert elements and are not chemically active.
2.2.4.4 Atoms with incompletely filled outer energy levels, like sodium and chlorine, tend to combine with other atoms in a chemical reaction. During the reaction, the atoms can trade off or share electrons and thereby fill their outer energy levels. Atoms that already have filled outer levels generally do not participate in chemical reactions for the simple reason that they do not need to gain or lose electrons. When two or more atoms combine in a chemical reaction, the resulting combination is called a molecule (MOL-e-kyool). A molecule may contain two atoms of the same kind, as in the hydrogen molecule: H2. The subscript 2 indicates that there are two hydrogen atoms in the molecule. Molecules may also be formed by the reaction of two or more different kinds of atoms, as in the hydrochloric acid molecule: HCL. Here an atom of hydrogen is attached to an atom of chlorine. A compound is a substance that can be broken down into two or more other substances by chemical means. The molecules of a compound always contain atoms of two or more different elements. Hydrochloric acid, which is present in the digestive juices of the stomach, is a compound. A molecule of hydrogen is not.
2.2.4.5 The atoms in a molecule are held together by electrical forces of attraction called chemical bonds, a form of potential energy. As you will see, when chemical bonds are broken, energy is released. When chemical bonds are formed, energy is required. Here we will consider ionic bonds, covalent bonds, and hydrogen bonds.
2.2.4.6 CLINICAL APPLICADON: lasers
2.2.4.6.1 The laser (acronym for light amplification by stimulated emission of radiation) is based on a relatively simple principle. Atoms, molecules, or ions in a laser are excited by absorption of energy (thermal, electrical, or optical). After absorption, the atoms, molecules, or ions give off an equal amount of energy in the form of light. This energy, the laser beam, is an intense light that can produce surgical effects. These include coagulation to stop bleeding, incision making, and tissue removal.
2.2.4.6.2 Clinical applications have been found for lasers in several medical and surgical specialties. Among these are ophthalmology (treatment of glaucoma, tumors, cataracts), dermatology and plastic surgery (removal of lesions, port-wine stains, and tattoos), cardiovascular surgery (angioplasty to open blood vessels), gastrointestinal surgery (control of bleeding and tumor removal), general surgery (incisions and removal of decubitus ulcers and leukoplakia), gynecology (treatment of vulvar, vaginal, and cervical neoplasia and endometriosis), neurosurgery (incision or removal of tissue near sensitive neural and vascular structures), otolaryngology (tonsillectomy, treatment of tumors of the larynx, and removal of nasal polyps), and urology (removal of lesions and opening strictures).
2.2.4.7 Ionic Bonds
2.2.4.7.1 Atoms are electrically neutral because the number of positively charged protons equals the number of negatively charged electrons. But when an atom gains or loses electrons, this balance is upset. If the atom gains electrons, it acquires an overall negative charge. If the atom loses electrons, it acquires an overall positive charge. Such a negatively or positively charged particle is called an ion (I-on).
2.2.4.7.2 Consider the sodium ion (Figure 2-3a). The sodium atom (Na) has 11 protons and 11 electrons, with 1 electron in its outer energy level. When sodium gives up the single electron in its outer level, it is left with 11 protons and only 10 electrons. It is an electron donor. The atom now has an overall positive charge of one (+ 1). This positively charged sodium atom is called a sodium ion (written Na+).
2.2.4.7.3 Another example is the formation of the chloride ion (Figure 2-3b). Chlorine has a total of 17 electrons, 7 of them in the outer energy level. Since this energy level can hold eight electrons, chlorine tends to pick up an electron that has been lost by another atom. Chlorine is an electron acceptor. By accepting an electron, chlorine acquires a total of 18 electrons. However, it still has only 17 protons in its nucleus. The chloride ion therefore has a negative charge of one ( - 1) and is written as Cl-.
2.2.4.7.4 The positively charged sodium ion (Na +) and the negatively charged chloride ion (Cl-) attract each other-unlike charges attract each other. The attraction, called an ionic bond, holds the two ions together, and a molecule is formed (Figure 2-3c). The formation of this molecule, sodium chloride (NaCl), or table salt, is one of the most common examples of ionic bonding. Thus, an ionic bond is an attraction between ions formed when one atom loses electrons and another atom gains electrons. Generally, atoms whose outer energy level is less than half filled lose electrons and form positively charged ions called cations (KAT-ī-ons). Examples of cations are potassium ion (K+), calcium ion (Ca2+), iron ion (Fe2+), and sodium ion (Na +). By contrast, atoms whose outer energy level is more than half-filled tend to gain electrons and form negatively charged ions called anions (AN- ī -ons). Examples of anions include iodide ion (I -), chloride ion (Cl-), and sulfur ion (S2-).
2.2.4.7.5 Notice that an ion is always symbolized by writing the chemical abbreviation followed by the number of positive ( + ) or negative ( - ) charges the ion acquires.
2.2.4.7.6 Hydrogen is an example of an atom whose outer level is exactly half-filled. The first energy level can hold two electrons, but in hydrogen atoms, it contains only one. Hydrogen may lose its electron and become a positive ion (H +). This is precisely what happens when hydrogen combines with chlorine to form hydrochloric acid (H + Cl-). However, hydrogen is equally capable of forming another kind of bond called a covalent bond.
2.2.4.8 Figure 2-3
2.2.4.8.1 Introduction
2.2.4.8.1.1 Formation of an ionic bond. (a) An atom of sodium attains stability by passing a single electron to an electron acceptor. The loss of this single electron results in the formation of a sodium ion (Na+). (b) An atom of chlorine attains stability by accepting a single electron from an electron donor. The gain of this single electron results in the formation of a chloride ion (GI-). (c) When the Na + and the CI- ions are combined, they are held together by the attraction of opposite charges, which is known as an ionic bond, and a molecule of sodium chloride (NaCI) is formed.
2.2.4.8.2 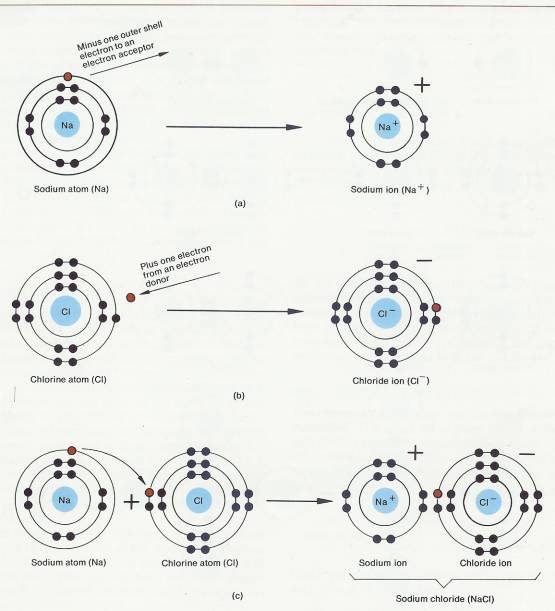
2.2.4.9 Covalent Bonds
2.2.4.9.1 The second chemical bond to be considered is the covalent bond. This bond is far more common in organisms than an ionic bond and is more stable. When a covalent bond is formed, neither of the combining atoms loses or gains an electron. Instead, the two atoms share one, two, or three electron pairs. Look at the hydrogen atom again. One way a hydrogen atom can fill its outer energy level is to combine with another hydrogen atom to form the molecule H2 (Figure 2-4a). In the H2 molecule, the two atoms share a pair of electrons. Each hydrogen atom has its own electron plus one electron from the other atom. The two electrons actually circle the nuclei of both atoms, spending their time equally between both atoms. Therefore, the outer energy levels of both atoms are filled half the time. When one pair of electrons is shared between atoms, as in the H2 molecule, a single covalent bond is formed. A single covalent bond is expressed as a single line between the atoms (H-H). When two pairs of electrons are shared between two atoms, a double covalent bond is formed, which is expressed as two parallel lines (=) (Figure 2-4b). A triple covalent bond, expressed by three parallel lines, occurs when three pairs of electrons are shared (Figure 2-4c).
2.2.4.9.2 The same principles that apply to covalent bonding between atoms of the same element also apply to atoms of different elements. Methane (CH4), also known as marsh gas, is an example of covalent bonding between atoms of different elements (Figure 2-4d). The outer energy level of the carbon atom can hold eight electrons but has only four of its own. Each hydrogen atom can hold two electrons but has only one of its own. In the methane molecule the carbon atom shares four pairs of electrons. One pair is shared with each hydrogen atom. Each of the four carbon electrons orbits around both the carbon nucleus and a hydrogen nucleus. Each hydrogen electron circles around its own nucleus and the carbon nucleus.
2.2.4.9.3 FIGURE 2-4 Covalent Bond
2.2.4.9.3.1 Introduction
2.2.4.9.3.1.1 Formation of a covalent bond between atoms of the same element and between atoms of different elements. In the symbols on the right, each covalent bond is represented by a straight line between atoms. (a) A single covalent bond between two hydrogen atoms. (b) A double covalent bond between two oxygen atoms. (c) A triple covalent bond between two nitrogen atoms. (d) Single covalent bonds between a carbon atom and four hydrogen atoms.
2.2.4.9.3.2 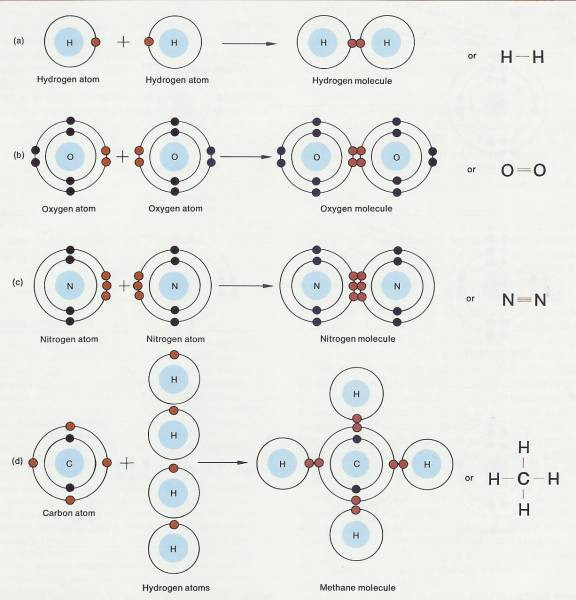
2.2.4.9.4 In some covalent bonds, the electrons are shared equally between atoms, that is, one atom does not attract the shared electrons more strongly than the other atom. Such covalent bonds are referred to as nonpolar covalent bonds. Examples include the bonds between hydrogen atoms and between oxygen atoms (see Figure 2-4a,b). In other covalent bonds, there is an unequal sharing of electrons between atoms, that is, one atom attracts the shared electrons more strongly than the other. This type of covalent bond is known as a polar covalent bond. An example of this type of bond occurs in a molecule of water (see Figure 2-6). This will be considered in more detail later in the chapter as part of the discussion of the function of water as a solvent.
2.2.4.9.5 Elements whose outer energy levels are half-filled, such as hydrogen and carbon, form covalent bonds quite easily. In fact, carbon always forms covalent bonds. It never becomes an ion. However, many atoms whose outer energy levels are more than half-filled also form covalent bonds. An example is oxygen. We won't go into the reasons why some atoms tend to form covalent bonds rather than ionic bonds.
2.2.4.9.6 Hydrogen Bonds
2.2.4.9.6.1 A hydrogen bond consists of a hydrogen atom covalently bonded to one oxygen atom or one nitrogen atom but attracted to another oxygen or nitrogen atom. Because hydrogen bonds are weak, only about 5 percent as strong as covalent bonds, they do not bind atoms into molecules. However, they do serve as bridges between different molecules or between various parts of the same molecule. The weak bonds may be formed and broken fairly easily. It is this property that accounts for the temporary bonding between certain atoms within large complex molecules such as proteins and nucleic acids (see Figure 2-12). It should be noted that even though hydrogen bonds are relatively weak, such large molecules might contain several hundred of these bonds, resulting in considerable strength and stability.
2.2.4.9.6.2 Chemical reactions are nothing more than the making, or breaking of bonds between atoms. And these reactions occur continually in all the cells of your body. As you will see again and again, chemical reactions are the processes by which body structures are built and body functions carried out.
2.2.5 Chemical Reactions
2.2.5.1 As we said earlier, chemical reactions involve the making or breaking of bonds between atoms. After a chemical reaction, the total number of atoms remains the same, but because they are rearranged, there are new molecules with new properties. In this section, we will look at the basic chemical reactions common to all living cells. Once you have learned them, you will be able to understand the chemical reactions discussed later.
2.2.5.2 Synthesis Reactions-Anabolism.
2.2.5.2.1 When two or more atoms, ions, or molecules combine to form new and larger molecules, the process is called a synthesis reaction. The word synthesis means "combination," and synthesis reactions involve the farming of new bonds. Synthesis reactions can be expressed in the following way:
2.2.5.2.2 Illustration Synthesis Reactions #1
2.2.5.2.2.1 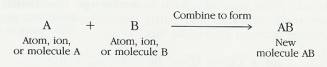
2.2.5.2.3 The combining substances, A and B, are called the reactants; the substance formed by the combination is the end product. The arrow indicates the direction in which the reaction is proceeding. An example of a synthesis reaction is:
2.2.5.2.4 Illustration Synthesis Reactions #2
2.2.5.2.4.1 
2.2.5.2.5 All the synthesis reactions that occur in your body are collectively called anabolic reactions, or simply anabolism (a-NAB-ō-lizm). Combining glucose molecules to form glycogen and combining amino acids to form proteins are two examples of anabolism. The importance of anabolism is considered in detail in Chapter 25.
2.2.5.3 Decomposition Reactions-Catabolism
2.2.5.3.1 The reverse of a synthesis reaction is a decomposition reaction. The word decompose means to break down into smaller parts. In a decomposition reaction, the bonds are broken. Large molecules are broken down into smaller molecules, ions, or atoms. A decomposition reaction occurs in this way:
2.2.5.3.2 Illustration Decomposition Reaction #1
2.2.5.3.2.1 
2.2.5.3.3 Under the proper conditions, methane can decompose into carbon and hydrogen:
2.2.5.3.4 Illustration Decomposition Reaction #2
2.2.5.3.4.1 
2.2.5.3.5 The subscript 4 on the left-hand side of the reaction equation indicates that four atoms of hydrogen are bonded to one carbon atom in the methane molecule. The number 4 on the right-hand side of the equation shows that four single hydrogen atoms have been set free.
2.2.5.3.6 All the decomposition reactions that occur in your body are collectively called catabolic reactions, or simply catabolism (ka-TAB- ō -lizm). The digestion and oxidation of food molecules are examples. The importance of catabolism is also considered in detail in later Chapters.
2.2.5.4 Exchange Reactions
2.2.5.4.1 All chemical reactions are based on synthesis or decomposition processes. In other words, chemical reactions are simply the making and/or breaking of ionic or covalent bonds. Many reactions, such as exchange reactions, are partly synthesis and partly decomposition. An exchange reaction works like this:
2.2.5.4.1.1 Illustration Exchange Reaction
2.2.5.4.1.1.1 
2.2.5.4.2 The bonds between A and B and between C and D are broken in a decomposition process. New bonds are then formed between A and D and between B and C or between A and C and between B and D in a synthesis process.
2.2.5.5 Reversible Reactions
2.2.5.5.1 When chemical reactions are reversible, the end product can revert to the original combining molecules. A reversible reaction is indicated by two arrows:
2.2.5.5.2 Illustration Reversible Reactions 1
2.2.5.5.2.1 
2.2.5.5.3 Some reversible reactions reverse themselves only under special conditions:
2.2.5.5.4 Illustration Reversible Reactions 2
2.2.5.5.4.1 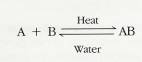
2.2.5.5.5 Whatever is written above or below the arrows indicates the special condition under which the reaction occurs. In this case, A and B react to produce AB only when heat is applied, and AB breaks down into A and B only when water is added. Figure 2-5 summarizes the basic chemical reactions that can occur.
2.2.5.6 How Chemical Reactions Occur
2.2.5.6.1 The collision theory explains how chemical reactions occur and how certain factors affect the rates of those reactions. According to this theory, all atoms, ions, and molecules are continuously moving and colliding with one another. The energy transferred by the particles in the collision might disrupt their electron structures enough that chemical bonds are broken or new ones are formed.
2.2.5.6.2 Several factors determine whether a collision will actually cause a chemical reaction. Among these are the velocities of the colliding particles, their energy, and their specific chemical configurations. Up to a point, the higher the particles' velocities, the greater the probability that their collision will result in a reaction. Also, each chemical reaction requires a specific level of energy. The collision energy required for a chemical reaction is its activation energy, which is the amount of energy needed to disrupt the stable electronic configuration of a specific molecule so that the electrons can be rearranged. (The relationship of enzymes to activation energy is considered in Chapter 25.) But even if colliding particles possess the minimum energy needed for reaction, no reaction will take place unless the particles are properly oriented toward each other.
2.2.5.7 Energy and Chemical Reactions
2.2.5.7.1 Energy is the capacity to do work. The two principal kinds of energy are potential (inactive or stored) and kinetic (energy of motion). Energy, whether potential or kinetic, exists in a number of different forms.
2.2.5.7.2 Chemical energy is the energy released or absorbed in the breaking or forming of chemical bonds. When a chemical bond is formed, energy is required. Such a chemical reaction is called an endergonic (energy inward) reaction. When a bond is broken, energy is released. Such a chemical reaction is called an exergonic (energy. outward) reaction. This means that synthesis reactions are endergonic (need energy), whereas decomposition reactions are exergonic (give off energy). The building processes of the body-the construction of bones, the growth of hair and nails, the replacement of injured cells occur basically through synthesis reactions. The breakdown of foods, on the other hand, occurs through decomposition reactions. When foods are decomposed, they release energy that can be used by the body for its building processes.
2.2.5.7.3 Radiant energy, such as heat and light, travels in waves. Some of the energy released during decomposition reactions is heat energy, which is used to help maintain normal body temperature, but does not accomplish cellular work.
2.2.5.7.4 Electrical energy is the result of the flow of charges, electrons, or charged particles called ions. As you will see later, electrical energy is essential for the conduction of action potentials by nerve and muscle cells.
2.2.5.7.5 The various forms of energy can be transduced (transformed) from one form into another. For example, potential energy can be transduced to kinetic energy and kinetic energy can be transduced to potential energy.
2.3 Chemical Compounds and Life Processes
2.3.1 Most of the chemicals in the body exist in the form of compounds. Biologists and chemists divide these compounds into two principal classes: inorganic compounds and organic compounds. Inorganic compounds usually lack carbon. They are usually small, ionically bonded molecules that are vital to body functions. They include water and many salts, acids, and bases. Organic compounds always contain carbon and hydrogen. Carbon is a unique element in the chemistry of life. Because carbon has four electrons in its outer shell, it can combine with a variety of atoms, including other carbon atoms, to form straight or branched chains and rings. Carbon chains are. the backbone for many substances of living cells. Organic compounds are held together mostly or entirely by covalent bonds. Organic compounds present in the body include carbohydrates, lipids, proteins, nucleic acids, and adenosine triphosphate (ATP).
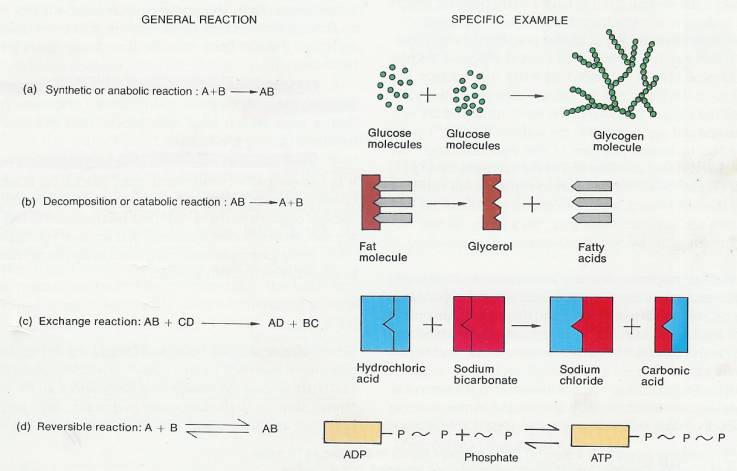
2.3.2 Figure 2-5
2.3.2.1 Description
2.3.2.1.1 FIGURE 2-5 Kinds of chemical reactions. (a) Synthetic or anabolic reaction. When linked together as shown, molecules of glucose form a molecule of glycogen. Glucose is a sugar that is the primary source of energy. Glycogen is a stored form of that sugar found in the liver and skeletal muscles. (b) Decomposition or catabolic reaction. The example shows a molecule of fat breaking down into glycerol and fatty acids. This reaction occurs whenever a food that contains fat is digested. (c) Exchange reaction. In this reaction, atoms of different molecules are exchanged with each other. Shown is a buffer reaction in which the body eliminates strong acids to help maintain homeostasis. (d) Reversible reaction. ATP (adenosine triphosphate) is an important source of stored energy. When such energy is needed, the ATP breaks down into ADP (adenosine diphosphate) and PO43- (phosphate group), releasing energy in the reaction. The phosphate group is symbolized as P. The cells of the body reconstruct ATP by using the energy of foods to attach ADP to PO43-
2.3.2.2
2.3.3 Inorganic Compounds
2.3.3.1 Water
2.3.3.1.1 One of the most important, as well as the most abundant, inorganic substances in the human organism is water. In fact, with a few exceptions, such as tooth enamel and bone tissue, water is by far the most abundant material in tissues. About 60 percent of red blood cells, 75 percent of muscle tissue, and 92 percent of blood plasma is water. The following functions of water explain why it is such a vital compound in living systems:
2.3.3.1.1.1 Water is an excellent solvent and suspending median. A solvent is a liquid or gas in which: some oilier material (solid, liquid, or gas), called a solute, has been dissolved. The combination of solvent plus solute is called a solution, and one common example of a solution is salt water. A solute, such as salt when placed in water, "'typically does not settle out of its solution. The solute can be retrieved through a chemical reaction or, in some cases, by boiling off the solvent. In a suspension, by contrast, the suspended material mixes with the liquid or suspending medium, but it will eventually settle out of the mixture. An example of a suspension is cornstarch and water. If the two materials are shaken together, a milky mixture forms. After the mixture sits for a while, however, the water clears at the top and the cornstarch settles to the bottom. Since water serves as a solvent for so many solutes, it is referred to as the universal solvent. .
2.3.3.1.1.2 The versatility of water as a solvent is related to its polar covalent bonds. Recall that a molecule of water contains polar covalent bonds, that is, bonds in which there is an unequal sharing of electrons between atoms. One atom attracts the shared electrons more strongly than others. In a molecule of water, there are both positive and negative areas (Figure 2-6a). When the two hydrogen atoms bond covalently to an oxygen atom, the shared electrons spend more time around oxygen than hydrogen. Since electrons have a negative charge, the unequal sharing causes the oxygen atom to have a slight negative charge and each hydrogen atom to have a slight positive charge.
2.3.3.1.1.3 In order to understand the solvating property of water, consider the following: If a crystal of an ionic compound, such as sodium chloride (NaCL), is placed in water, the sodium and chloride ions at the surface of the salt are exposed to the water molecules. The oxygen portions of the water molecule are negatively charged and are attracted to the sodium ion (Na +) of the salt, while the hydrogen portions of the water molecule are positively charged and are attracted to the chloride ions (Cl-) of the salt (Figure 2-6b). As the pattern repeats itself from the surface of the salt inward, water molecules surround the (Na +) and Cl- ions and separate them from each other. In this way, the salt is taken apart by water molecules-it is dissolved in water.
2.3.3.1.1.4 The solvating property of water is essential to health and survival. For example, if the surfaces of the air sacs in your lungs are not moist, oxygen cannot dissolve and therefore cannot move into your blood to be distributed throughout your body. Water, moreover, is "the solvent that carries nutrients into and wastes out of your body cells.
2.3.3.1.1.5 As a suspending medium, water is also vital to your survival. Many large organic molecules are suspended in the water of your body cells. These molecules are consequently able to come in contact with other chemicals, allowing various essential chemical reactions to occur.
2.3.3.1.1.6 Water can participate in chemical reactions
2.3.3.1.1.6.1 During digestion, for example, water can be added to large nutrient molecules in order to break them down into smaller molecules. This kind of breakdown is necessary if the body is to utilize the energy in nutrients. Water molecules are also used in synthesis reactions. Such reactions occur in the production of hormones and enzymes.
2.3.3.1.1.7 Water absorbs and releases heat very slowly
2.3.3.1.1.7.1 In comparison to other substances, water requires a large amount of heat to increase its temperature and a great loss of heat to decrease its. temperature. Thus, the presence of a large amount of water moderates the effects of fluctuations in environmental temperature and thereby helps to maintain a homeostatic body temperature.
2.3.3.1.1.8 Water requires a large amount of heat to change from a liquid to a gas
2.3.3.1.1.8.1 When water (perspiration) evaporates from the skin, it takes with it large quantities of heat and provides an excellent cooling mechanism.
2.3.3.1.1.9 Water serves as a lubricant in various regions of the body
2.3.3.1.1.9.1 It is a major part of mucus and other lubricating fluids. Lubrication is especially necessary in the chest and abdomen, where internal organs touch and slide over each other. It is also needed at joints, where bones, ligaments, and tendons rub against each other. In the gastrointestinal tract, water in mucus moistens foods to ensure their smooth passage
2.3.3.2 Figure 2-6
2.3.3.2.1 Solvating property of water. (a) Polar covalent bonds in a molecule of water. (b) The negative oxygen portions of the water molecules are attracted to the positive sodium ions (Na+), while the positive hydrogen portions of the water molecules-are attracted to the negative chloride ions (C-).
2.3.3.2.2 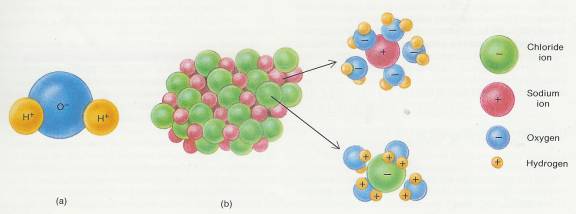
2.3.3.3 Inorganic Acids, Bases, and Salts
2.3.3.3.1 When molecules of inorganic acids, bases, or salts are dissolved in water in the body cells, they undergo ionization (ī-on-i-ZĀ-shun) or dissociation (dis'-sō-sē-Āshun); that is, they dissociate (separate) into ions. Such particles are also called electrolytes (ē-LEK-trō-līts) because the solution will conduct an electric current (the chemistry and importance of electrolytes are discussed in detail in Chapter 27). An acid may be defined as a substance that dissociates into one or more hydrogen ions (H+) and one or more negative ions (anions). Since an H + ion is a single proton with a charge of + 1, an acid may also be defined as a proton donor. A base, by contrast, dissociates into one or more hydroxyl ions (OH-) and one or more positive ions (cations). A base may also be viewed as a proton acceptor. Hydroxyl ions, as well as some other negative ions, have a strong attraction for protons. A salt, when dissolved in water, dissociates into cations and anions, neither of which is H+ or OH- (Figure 2-7). Acids and bases react with one another to form salts. For example, the combination of hydrochloric acid (HCI), an acid, and sodium hydroxide (NaOH), a base, produces sodium chloride (NaCl), a salt, and water (H2O ).
2.3.3.3.2 Many salts are found in the body. Some are in cells, whereas others are in the body fluids, such as lymph, blood, and the extracellular fluid of tissues. The ions, of salts are the source of many essential chemical elements. Exhibit 2-1 shows how salts dissociate into ions that provide these elements. Chemical analyses reveal that sodium and chloride ions are present in higher concentrations than other ions in extracellular body fluids. Inside the cells, phosphate and potassium ions are more abundant than other ions. Chemical elements such as sodium, phosphorus, potassium, or iodine are present in the body only in chemical combination with other elements or as ions. Their presence as free, un-ionized atoms could be instantly fatal. Exhibit 2-2 lists representative elements found in the body.
2.3.3.4 Exhibit 2-1
2.3.3.4.1 
2.3.3.5 Figure 2-7
2.3.3.5.1 Ionization of inorganic acids, bases, and salts. (a) When placed in water, hydrochloric acid (HCI) dissociates into H + ions and CI- ions. Acids are proton donors. (b) When the base sodium hydroxide (NaOH) is placed in water, it dissociates into OH- ions and Na+ ions. Bases are proton acceptors. (c) When table salt (NaCI) is placed in water, it dissociates into positive and negative ions (Na + and CI-), neither of which is H + or OH - .
2.3.3.5.2 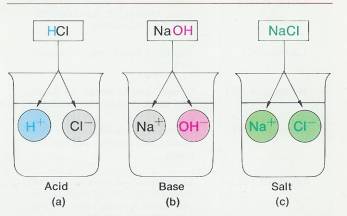
2.3.3.6 Acid-Base Balance: The Concept of pH
2.3.3.6.1 The fluids of your body must maintain a fairly constant balance of acids and bases. In solutions such as those found in body cells or in extracellular fluids, acids dissociate into hydrogen ions (H +) and anions. Bases, on the other hand, dissociate into hydroxyl ions (OH-) and cations. The more hydrogen ions that exist in a solution, the more acid the solution; conversely, the more hydroxyl ions, the more basic (alkaline) the solution. The term pH is used to describe the degree of acidity or alkalinity (basicity) of a solution.
2.3.3.6.2
Biochemical reactions
2.3.3.6.2.1
Reactions that occur in living systems-are extremely
sensitive to even small changes in the acidity or alkalinity of the environment
in which they occur. In fact, H+ and OH- ions are involved in practically all
biochemical processes, and the functions of cells are modified greatly by any
departure from narrow limits of normal H+ and OH- concentrations. For this
reason, the acids and bases that are constantly formed in the body must be kept
in balance. .
2.3.3.6.3 A solution's acidity or alkalinity is expressed on a pH scale that runs from 0 to 14 (Figure 2-8). The pH scale is based on the number of H+ ions in a solution expressed in chemical units called moles per liter. A pH of 7 means that a solution contains one ten-millionth (0.0000001) of a mole* of H+ ions per liter. The number 0.0000001 is written 10-7 in exponential form. To convert this value to pH, the negative exponent ( - 7) is converted into the positive number 7. A solution with a concentration of 0.0001 (10-4) of H+ ions per liter has a pH of 4; a solution with a concentration of 0.000000001 (10-9) has a pH of 9; and so on.
2.3.3.6.4 Exhibit 2-2
2.3.3.6.4.1 
2.3.3.6.5 A solution that is zero on the pH scale has many H+ ions and few OH - ions. A solution that rates 14, by contrast, has many OH- ions and few H+ ions. The midpoint in the scale is 7, where the concentration of H+ and OH ions is equal. A substance with a pH of 7, such as pure water, is neutral. A solution that has more H+ ions than OH- ions is an acid solution and has a pH below 7. A solution that has more OH- ions than H+ ions is a basic (alkaline) solution and has a pH above 7. A change of one whole number on, the pH scale represents a 10 fold change from the previous concentration; that is, a pH of 2 indicates 10 times fewer H + ions than a pH of 1. A pH of 3 indicates 10 times fewer H + ions than a pH of 2 and 100 times fewer H+ ions than a pH of 1.
2.3.3.6.6 * A mole of any substance is the weight, in grams, of the combined atomic weights of the atoms that make up a molecule of the substance. Example: A mole of H2O weighs 18 grams (2 for tl1e tWo hydrogen atoms + 16 for the oxygen atom).
2.3.3.7 Maintaining pH: Buffer Systems
2.3.3.7.1 Although the pH of body fluids may differ, the normal limits for the various fluids are generally quite specific and narrow. Exhibit 2-3 shows the pH values for certain body fluids compared with common substances. Even though strong acids and bases are continually taken into the body, the pH levels of these body fluids remain relatively constant. The mechanisms that maintain these homeostatic pH values in the body are called buffer systems. .
2.3.3.7.2 The essential function of a buffer system is to react with strong acids or bases in the body and replace them with weak acids or bases so that the strong acids or bases do not alter pH drastically. Strong acids (or bases) ionize easily and contribute many H+ (or OH-) ions to a solution. They therefore change the pH drastically. Weak acids (or bases) do not ionize so easily. They contribute fewer H + (or OH -) ions and have little effect on the pH. The chemicals that replace strong acids or bases with weak ones are called buffers and are found in the body's fluids. Most buffers in the human body consist of a weak acid and the salt of that acid. It is very important to note that the salt of the acid functions as a weak base. At this point, we will examine the carbonic acid--bicarbonate buffer system-the most important one found in extracellular fluid.
2.3.3.7.3 Figure 2-8
2.3.3.7.3.1 FIGURE 2-8 pH scale. At pH 7 (neutrality), the concentration of H+ and OH- ions is equal. A pH value below 7 indicates an acid solution; that is, there are more H+ ions than OH- ions. The lower the numerical value of the pH, the more acid the solution is because the H+ ion concentration becomes progressively greater. A pH value above 7 indicates an alkaline (basic) solution; that is, there are more OH- ions than H+ ions. The higher the numerical value of the pH, the more alkaline the solution is because the OH- ion concentration becomes progressively greater. A change in one whole number on the pH scale represents a 10-fold change from the previous concentration (100 = 1.0,10-1 = 0.1. 10-2 = 0.01. 10-3 = 0.001. and so on).
2.3.3.7.3.2 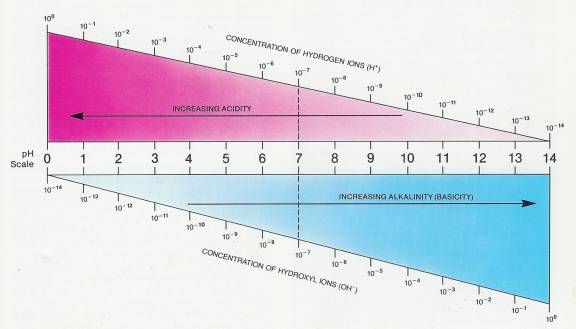
2.3.3.7.4 The carbonic acid-bicarbonate buffer system consists of a pair of chemicals: carbonic acid (H2CO3), which functions as the weak acid, and sodium bicarbonate (NaHCO3), the salt of the weak acid, which functions as the weak base. (As you will see in Chapter 27, it is actually the anion of the salt that behaves like the weak base.) The carbonic acid is a proton donor, that is, a hydrogen ion (H +) donor, and the bicarbonate ion (HCO3 -) of the sodium bicarbonate is a proton acceptor, that is, a hydrogen ion (H +) acceptor. In solution, the members of this buffer pair dissociate as follows:
2.3.3.7.5 Illustration Weak Acid Component & Salt of the Acid Component
2.3.3.7.5.1 
2.3.3.7.6 Each member of the buffer pair has a specific role in helping the body maintain a constant pH. If the body's pH is threatened by the presence of a strong acid, the salt of the acid of the buffer pair behaves like a weak base and goes into operation. If the body's pH is threatened by a strong base, the weak acid goes into play.
2.3.3.7.7 Consider the following situation. If a strong acid, such as HCI, is added to extracellular fluid, the salt of the acid of the buffer system behaves like a weak base and goes to work, and the following acid-buffering reaction occurs:
2.3.3.7.8 HCI Hydrochloric Acid
2.3.3.7.8.1 
2.3.3.7.9 The chloride ion (Cl-) of HCI and the sodium ion (Na +) of sodium bicarbonate combine to form NaCl, a substance that has no effect on pH. The hydrogen ion of the HCI could greatly lower pH by making the solution more acid, but this H + ion combines with the bicarbonate ion (HCO3 -) of sodium bicarbonate to form carbonic acid, a weak acid that lowers pH only slightly. In other words, because of the action of the salt of the acid of the buffer system functioning as a weak base, the strong acid (HCI) has been replaced by a weak acid (H2CO3) and a salt (NaCl), and the pH remains relatively constant.
2.3.3.7.10 Now suppose a strong base, such as sodium hydroxide (NaOH), is added to the extracellular fluid. In this instance, carbonic acid, the weak acid of the buffer system, goes to work and the following base-buffering reaction takes place
2.3.3.7.11 Exhibit 2-3
2.3.3.7.11.1

2.3.3.7.12 Illustration Base-Buffering Reaction
2.3.3.7.12.1

2.3.3.7.13 In this reaction, the OH- ion of sodium hydroxide could greatly raise the pH of the solution by making it more alkaline. However, the OH- ion combines with an H+ ion of carbonic acid and forms water, a substance that has no effect on pH. In addition, the Na + ion of sodium hydroxide combines with the bicarbonate ion (HCO3 -) to form sodium bicarbonate, a salt of the acid that acts like a weak base and has little effect on pH. Thus, because of the action of the buffer system, the strong base (NaOH) is replaced by water and a salt of the acid (NaHCO3), that functions like a weak base, and the pH remains relatively constant.
2.3.3.7.14 Whenever a buffering reaction occurs, the concentration of one member of the buffer pair is increased, whereas the concentration of the other decreases. When a strong acid is buffered, for example, the concentration of carbonic acid is increased, but the concentration of sodium bicarbonate is decreased. This happens because carbonic acid is produced and sodium bicarbonate is used up in the acid-buffering reaction. When a strong base is buffered, the concentration of sodium bicarbonate is increased, but the concentration of carbonic acid is decreased because sodium bicarbonate is produced and carbonic acid is used up in the base-buffering reaction. When the buffered substances-HCI and NaOH, in this case-are removed from the body via the kidneys, the carbonic acid and sodium bicarbonate formed as products of the reactions function again as components of the buffer pair. Do you now understand why buffers are sometimes called "chemical sponges"?
2.3.3.7.15 Although the preceding discussion of pH has focused on inorganic acids and bases, you should know that there are also organic acids and bases and that they, too, are involved in counteracting potential pH problems
2.3.4 Organic Compounds
2.3.4.1 In addition to carbon, the most frequently found elements in organic compounds are hydrogen (which can form one bond), oxygen (two bonds), and nitrogen (three bonds). Sulfur (two bonds) and phosphorus (five bonds) appear less often. Other elements are found, but only in a relatively few organic compounds. Carbon has several properties that make it particularly useful to living organisms. For one thing, it can react with one to several hundred other carbon atoms to form large molecules of many different shapes. This means that the body can build many compounds out of carbon, hydrogen, and oxygen. Each compound can be especially suited for a particular structure or function. The relatively large size of most carbon containing molecules and the fact that some do not dissolve easily in water make them useful materials for building body structures. Carron compounds are mostly or entirely held together by covalent bonds and tend to decompose easily. This means that organic compounds are also a good source of energy. Ionic compounds are not good energy sources because they form new ionic bonds as soon as the old ones are broken.
2.3.4.2 Carbohydrates
2.3.4.2.1 A large and diverse group of organic compounds found in the body are the carbohydrates, also known as sugars and starches. The carbohydrates perform a number of major functions in living systems. A few even form structural units. For instance, one type of sugar (deoxyribose) is a building block of deoxyribonucleic acid (DNA),'the molecule that carries hereditary information. Some carbohydrates are converted to other substances, which are used to build structures and provide an emergency source of energy. Other carbohydrates function as food reserves. One example is glycogen, which is stored in the liver and skeletal muscles. The principal function of carbohydrates, however, is to provide the most readily available source of energy to sustain life.
2.3.4.2.2 Carbon, hydrogen, and oxygen are the elements found in carbohydrates. The ratio of hydrogen to oxygen atoms is typically 2: 1, the same as in water. This ratio can be seen in the formulas for carbohydrates such as ribose (CH5H10O5), glucose (C6H12O6), and sucrose (C12H22011). Although there are exceptions, the general formula for carbohydrates is (CH2O)n, where n symbolizes three or more CH2O units. Carbohydrates can be divided into three major groups on the basis of size: monosaccharides, disaccharides, and polysaccharides.
2.3.4.2.2.1 Monosaccharides
2.3.4.2.2.1.1
Monosaccharides
(mon-ō-SAKa-rīds), or simple
sugars, are compounds containing from three to seven carbon atoms. Simple
sugars with three carbons in the molecule are called trioses. The number of
carbon atoms in the molecule is indicated by the prefix tri. There are
also tetroses (four-carbon sugars), pentoses (five-carbon sugars), hexoses
(six-carbon sugars), and heptoses (seven-carbon sugars). Pentoses and hexoses
are exceedingly important to the human organism. The pentose called
deoxyribose is a component of genes.
The hexose called glucose is the main energy-supplying molecule of the
body.
2.3.4.2.2.2 Disaccharides
2.3.4.2.2.2.1
A second group
of carbohydrates, the disaccharides (dī-SAK-a-rīds),
are also sugars and consist of two monosaccharides joined chemically. In the
process of disaccharide formation, two monosaccharides combine to form a
disaccharide molecule and a molecule of water is lost. This reaction is known
as dehydration synthesis (dehydration = loss
of water). The following reaction shows disaccharide formation. Molecules of
the monosaccharides glucose and fructose combine to form a molecule of the
disaccharide sucrose (table sugar):
2.3.4.2.2.2.2
Illustration
Disaccharide Formation
2.3.4.2.2.2.2.1
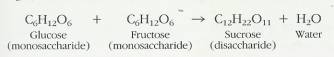
2.3.4.2.2.2.3 You may be puzzled to see that glucose and fructose have the same chemical formulas. Actually, they are different monosaccharides, since the relative positions of the oxygens and carbons vary in the two different molecules (see Figure 2-9). The formula for sucrose is C12H22011 and not C12H24012, since a molecule of H2O is lost in the process of disaccharide formation. In every dehydration synthesis, a molecule of water is lost. Along with this water loss, there is the synthesis of two small molecules, such as glucose and fructose, into one large, more complex molecule, such as sucrose (Figure 2-9). Similarly, the. dehydration synthesis of the two monosaccharides glucose and galactose forms the disaccharide lactose (milk sugar).
2.3.4.2.2.2.4 Disaccharides can also be broken down into smaller, simpler molecules by adding water. This reverse chemical reaction is called digestion (hydrolysis), which means to split by using water. A molecule of sucrose, for example, may be digested into its components of glucose and fructose by the addition of water. The mechanism of this reaction also is represented in Figure 2-9,
2.3.4.2.2.3 Polysaccharides
2.3.4.2.2.3.1 The third major group of carbohydrates, the polysaccharides (pol' -ē-SAK-a-rīds), consists of several monosaccharides joined together through dehydration synthesis. Polysaccharides have the formula (C6HIOO5)n. Like disaccharides, polysaccharides can be broken down into their constituent sugars through hydrolysis reactions. Unlike monosaccharides or disaccharides, however, they usually lack the characteristic sweetness of sugars like fructose or sucrose and are usually not soluble in water. One of the chief polysaccharides is glycogen.
2.3.4.2.3 Clinical Applications: Artificial Sweeteners
2.3.4.2.3.1 
2.3.4.2.4 Figure 2-9
2.3.4.2.4.1 FIGURE 2-9 Dehydration synthesis and hydrolysis of a molecule of sucrose. In the dehydration synthesis reaction (read from left to right), the two smaller molecules, glucose and fructose, are joined to form a larger molecule of sucrose. Note the loss of a water molecule. In hydrolysis (read from right to left), the larger sucrose molecule is broken down into the two smaller molecules, glucose and fructose. Here, a molecule of water is added to sucrose for the reaction to occur.
2.3.4.2.4.2 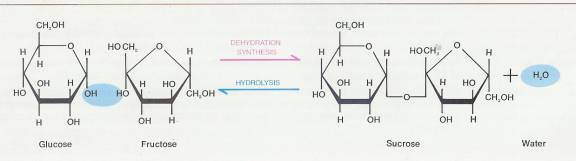
2.3.4.2.4.3
2.3.4.3 Lipids
2.3.4.3.1 A second group of organic compounds that is vital to the human organism is the lipids. Like carbohydrates, lipids are composed of carbon, hydrogen, and oxygen, but they do not have a 2:1 ratio of hydrogen to oxygen. In fact, the amount of oxygen in lipids is usually less than that in carbohydrates. Most lipids are insoluble in water, but they readily dissolve in solvents such as alcohol, chloroform, and ether. Among the groups of lipids are fats, phospholipids (lipids that contain phosphorous), steroids, carotenes, vitamins E and K, and prostaglandins (PGs). A listing of the various types of lipids is shown in Exhibit 2-4, along with their relationships to the human organism. Since lipids are a large and diverse group of compounds, we will discuss only two types in detail at this point: fats and prostaglandins.
2.3.4.3.2 A molecule of fat (triglyceride) consists of two basic components: glycerol and fatty acids (Figure 2-10). A single molecule of fat is formed when a molecule of glycerol combines with three molecules of fatty acids. This reaction, like the one described for disaccharide formation, is a dehydration synthesis reaction. During hydrolysis, a single molecule of fat is broken down into fatty acids and glycerol.
2.3.4.3.3 Exhibit 2-4
2.3.4.3.3.1 
2.3.4.3.4 In subsequent chapters, we will be talking about saturated, monosaturated, and polyunsaturated fats. These terms have the following meanings. A saturated fat contains no double bonds between any of its carbon atoms. It contains only single covalent bonds between its carbon atoms, and all the carbon atoms are bonded to the maximum number of hydrogen atoms; thus, such a fat is saturated with hydrogen atoms (Figure 2-10c). Saturated fats (and some cholesterol) occur mostly in animal foods such as beef, pork, butter, whole milk, eggs, and cheese. They also occur in some plant products such as cocoa butter, palm oil, and coconut oil. Since the liver uses some breakdown products of saturated fats to produce cholesterol, consumption of these fats is discouraged for individuals with high cholesterol levels. A monosaturated fat contains one double covalent bond between its carbon atoms; it is not completely saturated with hydrogen atoms (Figure 2-10c). Examples are olive oil and peanut oil, which are believed to help reduce cholesterol levels. A polyunsaturated fat contains more than one double covalent bond between its carbon atoms (Figure 2-10c). Corn oil, safflower oil, sunflower oil, cottonseed oil, sesame oil, and soybean oil are examples of polyunsaturated fats, which researcher believe also help to reduce cholesterol in the blood.
2.3.4.3.5 Fats represent the body’s most highly concentrated source of energy.
2.3.4.3.6 They provide more than twice as much energy per-weight as either carbohydrates or proteins. In addition, fats do not attract water and thus do not result in excessive water retention in the body. In general, however, fats are about 10 to 12 percent less efficient as body fuels than are carbohydrates. A great amount of the fat calorie is wasted and thus not available for the body to use.
2.3.4.3.7 FIGURE 2-10
2.3.4.3.7.1 Structure and reactions of glycerol and fatty acids. (a) Structure of glycerol. (b) Structure of a fatty acid. The one shown here is palmitic acid. Note that when glycerol and the fatty acid are joined in dehydration synthesis, a molecule of water is lost. (c) Fats consist of one molecule of glycerol joined to three molecules of fatty acids, which vary in length and the number and location of double bonds between carbon atoms (C=C). Shown here is a molecule of a fat that contains three different fatty acids: palmitic acid, a saturated fatty acid; lineolic acid, a polyunsaturated fatty acid; and oleic acid, a monosaturated fatty acid. Note that when a molecule of glycerol combines with three fatty acids in dehydration synthesis to form a molecule of fat, three molecules of water are lost.
2.3.4.3.7.2
2.3.4.3.8 
2.3.4.3.9 Clinical Application
2.3.4.3.9.1 
2.3.4.3.10 Prostaglandins (pros'-ta-GLAN-dins), also called PGs, are a large group of membrane-associated lipids composed of 20-carbon fatty acids containing 5 carbon atoms joined to form a ring (cyclopentane ring). Prostaglandins are produced in all nucleated cells in the body and are able to influence the functioning of any type of cell.
2.3.4.3.11 Prostaglandins are produced in cell membranes and are rapidly decomposed by catabolic enzymes. Although synthesized in minute quantities, they are potent substances and exhibit a wide variety of effects on the body. Basically, prostaglandins mimic hormones. They are involved in modulating many hormonal responses (Chapter 18), inducing menstruation, and inducing second-trimester abortions. They are also involved in contributing to the inflammatory response (Chapter 22), preventing peptic ulcers, opening bronchial and nasal passages, platelet aggregation and inhibition of aggregation, and regulating body temperature.
2.3.4.4 Proteins
2.3.4.4.1 A third principal group of organic compounds is proteins: These compounds are much more complex in structure than the carbohydrates or lipids. They are also responsible for much of the structure of body cells and are related to many physiological activities. For example, proteins in the form of enzymes speed up most essential biochemical reactions. This is described in detail in Chapter 25. Other proteins assume a necessary role in muscular contraction. Antibodies are proteins that provide the human organism with defenses against invading microbes. And some hormones that regulate body functions are also proteins. A classification of proteins on the basis of function is shown in Exhibit 2-5.
2.3.4.4.2 Chemically, proteins always contain carbon, hydrogen, oxygen, and nitrogen. Many proteins also contain sulfur and phosphorus. Just as monosaccharides are the building units of sugars, and fatty acids and glycerol are the building units of fats, amino acids are the building blocks of proteins. In protein formation, amino acids combine to form more complex molecules, while water molecules are lost. The process is a dehydration synthesis reaction, and the bonds formed between amino acids are called peptide bonds (Figure 2-11).
2.3.4.4.3
When two amino acids combine, a dipeptide results. Adding another
amino acid to a dipeptide produces a tripeptide.
Further additions of amino acids result in the formation of chainlike polypeptides, which are large
protein molecules that consist of 50 or more amino acids. At least 20 different
amino acids are found in proteins. Although all amino acids have one component
in common, each is distinguished on the basis of additional atoms or groups of
atoms arranged in a specific way. A great variety of proteins are possible
because each variation in the number or sequence of amino acids can produce a
different protein. The situation is similar to using an alphabet of 20 letters
to form words. Each letter could be compared to a different amino acid, and each
word would be a different protein.
2.3.4.4.4 Exhibit 2-5
2.3.4.4.4.1 
2.3.4.4.5 Figure 2-11
2.3.4.4.5.1 Protein formation. When two or more amino acids are chemically united, the resulting bond between them is called a peptide bond. In the example shown here, the amino acids glycine and alanine are joined to form the dipeptide glycylalanine. The peptide bond is formed at the point where water is lost.
2.3.4.4.5.2 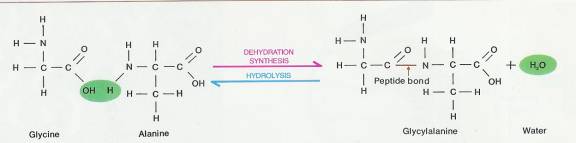
2.3.4.4.6 Proteins vary tremendously in structure. Different proteins have different architectures and different three-dimensional shapes. This variation in structure and shape is directly related to their diverse functions. When a cell makes a protein, the polypeptide chain folds spontaneously to assume a certain shape. One reason for folding of the polypeptide is that some parts of a protein are attracted to water, and other parts are repelled by it. In practically every case, the function of a protein depends on its ability to recognize and bind to some other molecule, a situation similar to the fit between a lock and key; As an example, an enzyme binds specifically with its substrate. A hormonal protein binds to a receptor on a cell whose function it will alter. An antibody binds to a foreign substance (antigen) that has invaded the body. The unique shape of a protein permits it to interact with other specific molecules in order to carry out specific functions.
2.3.4.4.7 Proteins exhibit four levels of structural organization. The primary structure is the unique order (sequence) of amino acids making up the protein. An alteration in primary structure can have serious consequences. For example, a single substitution of an amino acid in a blood protein can result in a deformed hemoglobin molecule that produces sickle-cell anemia. The secondary structure of a protein is the localized, repetitious twisting or folding of its polypeptide chain. Common secondary structures are clockwise spirals (helixes) and pleated sheets. The tertiary structure refers to the overall three-dimensional structure of a polypeptide chain. The folding is not repetitive or predictable as in secondary structures. The tertiary structure is very irregular, and it determines the particular function of a protein. The quaternary structure of a protein refers to two or more individual polypeptide chains bonded to each other that function as a single unit.
2.3.4.4.8
It was once thought that the atoms in proteins are in
fixed positions and that proteins are therefore rigid molecules. However,
based on computer simulations, it has been learned that the atoms in proteins
are in constant motion, usually twisting movements. Such motion is due to the
chemical bonds between atoms that act like springs. The internal motion of
proteins results in their constantly changing structure and explains the many
and varied functions of proteins. Scientists are now studying how proteins
fold into three-dimensional structures that determine their biological
functions.
2.3.4.4.9 If a protein encounters a hostile environment in terms of temperature, pH, or salt concentrations, it may unravel and lose its characteristic shape. This process is called denaturation. As a result of denaturation, the protein is no longer functional.
2.3.4.4.10 Nucleic Acids: Deoxyribonucleic Acid (DNA) and Ribonucleic Acid (RNA)
2.3.4.4.10.1 Nucleic (noo-KLĒ-ic) acids, compounds first discovered in the nuclei of cells, are exceedingly large organic molecules containing carbon, hydrogen, oxygen, nitrogen, and phosphorus. They are divided into two principal kinds: deoxyribonucleic (dē-ok' -sē-rī' -bō-noo-KLĒ-ik) acid (DNA) ,:and ribonucleic acid (RNA). I
2.3.4.4.10.2 Whereas the basic structural units of proteins are amino acids, the basic units of nucleic acids are nucleotides. A molecule of DNA is a chain composed of repeating nucleotide units. Each nucleotide of DNA consists of three basic parts (Figure 2-12a):
2.3.4.4.10.2.1
It contains
one of four possible nitrogenous bases, which are ring-shaped structures
containing atoms of C, H, O, and N. The nitrogenous bases found in DNA are
adenine, Thymine, cytosine, and guanine. Adenine and guanine are double-ring
structures, collectively referred to as purines. Thymine and cytosine
are smaller, single-ring structures called pyrimidines.
2.3.4.4.10.2.2
It contains
a pentose sugar called deoxyribose.
2.3.4.4.10.2.3
It also
contains phosphate groups.
2.3.4.4.10.3 The nucleotides are named according to the nitrogenous base that is present. Thus, a nucleotide containing thymine is called a thymine nucleotide. One containing adenine is called an adenine nucleotide, and so on.
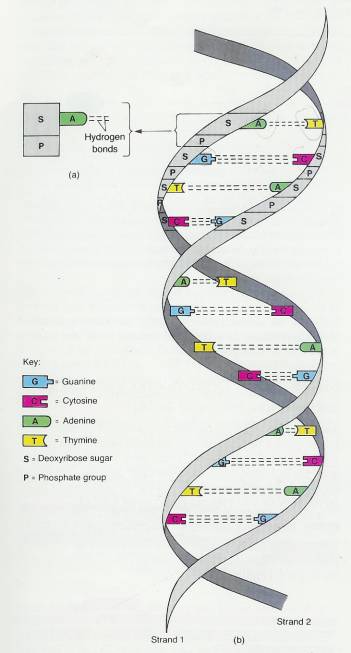
2.3.4.4.10.4 The chemical components of the DNA molecule were known before 1900, but it was not until 1953 that a model of the organization of the chemicals was constructed. This model was proposed by J. D. Watson and F. H. C. Crick on the basis of data from many investigations. Figure 2-12b shows the following structural characteristics of the DNA molecule.
2.3.4.4.10.4.1 The molecule consists of two strands with crossbars. The strands twist about each other in the form of a double helix, so that the shape resembles a twisted ladder. For many years, it was assumed that all DNA was in the form of a double helix that twisted smoothly to the right (right-handed DNA). There at least four varieties of right-handed DNA designated as A-DNA, B-DNA, C-DNA, and D-DNA. Watson and Crick's model, the one most commonly found in cells, is B-DNA. It was discovered, however, that DNA can also twist jaggedly to the left (Z-DNA or left-handed DNA). This alternate form of DNA is located near the ends of genes and may help to explain how genes turn on and off and how some cells may become malignant.
2.3.4.4.10.4.2 The uprights of the DNA ladder consist of alternating phosphate groups and the deoxyribose portions of the nucleotides.
2.3.4.4.10.4.3 . The rungs ofthe ladder contain paired nitrogenous bases. As shown, adenine always pairs with thymine, and cytosine always pairs with, guanine.
2.3.4.4.10.5 Figure 2-12
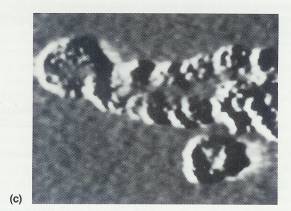
2.3.4.4.10.5.1 DNA molecule. (a) Adenine nucleotide. (b) Portion of an assembled DNA molecule. (c) Strand of DNA viewed through a scanning tunneling microscope. This is the first direct image of chemically unaltered, uncoated, pure DNA. The looped DNA strand stretches across an area 400 A wide. The circular structure is possibly an unresolved DNA fragment. (Courtesy of Lawrence Livermore National Laboratory.)
2.3.4.4.10.5.2
2.3.4.4.10.5.3
2.3.4.4.10.6 Cells contain hereditary material called genes, each of which is a segment of a DNA molecule. Our genes determine which traits we inherit, and they control all the activities that take place in our cells throughout a lifetime. When a cell divides, its hereditary information is passed on to the next generation of cells. The passing of information is possible because of DNA's unique structure.
2.3.4.4.10.7 Clinical Application
2.3.4.4.10.7.1

2.3.4.4.10.8 RNA, the second principal kind of nucleic acid, differs from DNA in several respects. RNA is single-stranded; DNA is double-stranded. The sugar in the RNA nucleotide is the pentose ribose. And RNA does not contain the nitrogen base thymine. Instead of thymine, RNA has the nitrogen base uracil. At least three different kinds of RNA have been identified in cells. Each type has a specific role to perform with DNA in protein synthesis reactions (Chapter 3).
2.3.4.5 Adenosine Triphosphate (ATP)
2.3.4.5.1 Figure 2-13
2.3.4.5.1.1 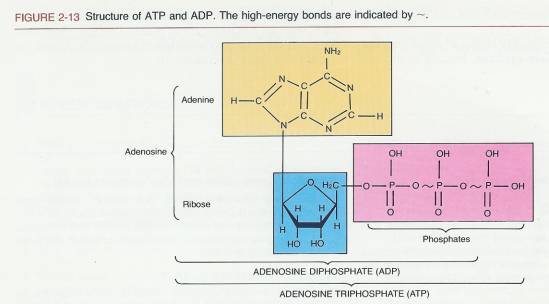
2.3.4.5.2 A molecule that is indispensable to the life of the cell is adenosine (a-DEN-o-sen) triphosphate (ATP). This substance is found universally in living systems and performs the essential function of storing energy for various cellular activities. Structurally, ATP consists of three phosphate groups (PO43-) and an adenosine unit composed of adenine and the five-carbon sugar ribose (Figure 2-13). ATP is regarded as a high-energy molecule because of the total amount of usable energy it releases when it is broken down by the addition of a water molecule (hydrolysis ).
2.3.4.5.3 When the terminal phosphate group (here symbolized by P) is hydrolyzed, the reaction liberates a great deal of energy. This energy is used by the cell to perform its basic activities. Removal of the terminal phosphate group leaves a molecule called adenosine diphosphate (ADP). This reaction may be represented as follows:
2.3.4.5.4 Illustration ADP
2.3.4.5.4.1 
2.3.4.5.5 The energy supplied by the catabolism of ATP into ADP is constantly being used by the cell. Since the supply of ATP at any given time is limited, a mechanism exists to replenish it-a phosphate group is added to ADP to manufacture more ATP. The reaction may be represented as follows:
2.3.4.5.6 Illustration ATP
2.3.4.5.6.1 
2.3.4.5.7 Logically, energy is required to manufacture ATP. The energy required to attach a phosphate group to ADP is supplied by various decomposition reactions taking place in the cell, particularly by the decomposition of glucose. Glucose is completely metabolized in cells into carbon dioxide and water, and the energy released in this process is used to attach a phosphate group to ADP to resynthesize ATP. ATP can be stored in every cell, where it provides potential energy that is not released until needed.
2.3.4.6 Cyclic AMP
2.3.4.6.1 A chemical substance closely related to ATP is cyclic AMP, also known as adenosine-3',5'-monophosphate. Essentially it is a molecule of adenosine monophosphate with the phosphate attached to the ribose sugar at two places (Figure 2-14). START
2.3.4.6.2 The attachment forms a ring-shaped structure and thus the name cyclic AMP.
2.3.4.6.3 Cyclic AMP is formed from ATP by the action of a special enzyme, called adenylate cyclase, located in the cell membrane. Although cyclic AMP was discovered in 1958, only recently has its function in cells become clear. One function is related to the action of hormones, a topic we explore in detail in Chapter 18.
2.3.4.6.4 Figure 2-14
2.3.4.6.4.1 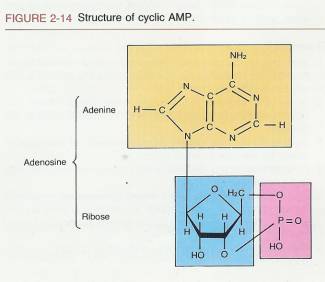
2.4 Summary
2.4.1 Introduction to Basic Chemistry (Link)
2.4.1.1 Chemical Elements (Link)
2.4.1.1.1 Matter is anything that occupies space and has mass. It is made up of building units called chemical elements.
2.4.1.1.2 Oxygen, carbon, hydrogen, and nitrogen make up 96 percent of body weight. These elements together with calcium and phosphorus make up 99 percent of total body weight.
2.4.1.2 Structure of Atoms (Link)
2.4.1.2.1 Units of matter of all chemical elements are called atoms.
2.4.1.2.2 Atoms consist of a nucleus, which contains protons and neutrons (nucleons), and electrons that move about the nucleus in energy levels.
2.4.1.2.3 The total number of protons of an atom is its atomic number. This number is equal to the number of electrons in the atom.
2.4.1.3 Atoms and Molecules (Link)
2.4.1.3.1 The electrons are the part of an atom that actively participate in chemical reactions.
2.4.1.3.2 A molecule is the smallest unit of two or more combined atoms. A molecule containing two or more different kinds of atoms is a compound.
2.4.1.3.3 In an ionic bond, outer-energy-level electrons are transferred from one atom to another. The transfer forms ions, whose unlike charges attract each other and form ionic bonds.
2.4.1.3.4 In a covalent bond, there is a sharing of pairs of outer-energy level electrons.
2.4.1.3.5 Hydrogen bonding provides temporary bonding between certain atoms within large complex molecules such as proteins and nucleic acids.
2.4.1.4 Chemical Reactions (Link)
2.4.1.4.1 Synthesis reactions involve the combination of reactants to produce a new molecule. The reactions are anabolic: bonds are formed.
2.4.1.4.2 In decomposition reactions, a substance breaks down into other substances. The reactions are catabolic: bonds are broken.
2.4.1.4.3 Exchange reactions involve the replacement of one atom or atoms by another atom or atoms.
2.4.1.4.4 In reversible reactions, end products can revert to the original combining molecules.
2.4.1.4.5 When chemical bonds are formed (endergonic reaction), energy is needed. When bonds are broken (exergonic reaction), energy is released. This is known as chemical bond energy.
2.4.1.4.6 Other forms of energy include mechanical, radiant, and electrical.
2.4.2 Chemical Compounds and Life Processes (Link)
2.4.2.1 Inorganic substances usually lack carbon, contain ionic bonds, resist decomposition, and dissolve readily in water.
2.4.2.2 Organic substances always contain carbon and hydrogen. Most organic substances contain covalent bonds and many are insoluble in water.
2.4.2.3 Inorganic Compounds (p. 37)
2.4.2.3.1 Water is the most abundant substance in the body. It is an excellent solvent and suspending medium, participates in chemical reactions, absorbs and releases heat slowly, and lubricates.
2.4.2.3.2 Inorganic acids, bases, and salts dissociate into ions in water. An acid ionizes into H+ ions; a base ionizes into OH- ions. A salt ionizes into neither H+ nor OH- ions. Cations are positively charged ions; anions are negatively charged ions.
2.4.2.3.3 The pH of different parts of the body must remain fairly constant for the body to remain healthy. On the pH scale, 7 represents neutrality. Values below 7 indicate acid solutions, and values above 7 indicate alkaline solutions.
2.4.2.3.4 The pH values of different parts of the body are maintained by buffer systems, which usually consist of a weak acid and a weak base. Buffer systems eliminate excess H+ ions and excess OH- ions in order to maintain pH homeostasis.
2.4.2.4 Organic Compounds (Link)
2.4.2.4.1 Carbohydrates are sugars or starches that provide most of the energy needed for life: They may be monosaccharides, disaccharides, or polysaccharides. Carbohydrates, and other organic molecules, are joined together to form larger molecules with the loss of water by a process called dehydration synthesis. In the reverse process, called digestion (hydrolysis), large molecules are broken down into smaller ones upon the addition of water.
2.4.2.4.2 Lipids are a diverse group of compounds that includes fats, phospholipids, steroids, carotenes, vitamins E and K, and prostaglandins (PGs). Fats protect, insulate, provide energy, and are stored. Prostaglandins mimic the effects of hormones and are involved in the inflammatory response and the modulation of hormonal responses.
2.4.2.4.3 Proteins are constructed from amino acids. They give structure to the body, regulate processes, provide protection,; help muscles to contract, transport substances, and serve as enzymes. Structural levels of organization among proteins include: primary, secondary, tertiary, and quaternary.
2.4.2.4.4 Deoxyribonucleic acid (DNA) and ribonucleic acid (RNA) are nucleic acids consisting of nitrogenous bases, sugar, and phosphate groups. DNA is a double helix and is the primary chemical in genes. RNA differs in structure and chemical composition from DNA and is mainly concerned with protein synthesis reactions.
2.4.2.4.5 The principal energy-storing molecule in the body is adenosine triphosphate (ATP). When its energy is liberated, it is decomposed to adenosine diphosphate (ADP) and P. ATP is manufactured from ADP and P using the energy supplied by various decomposition reactions, particularly of glucose.
2.4.2.4.6 . Cyclic AMP is closely related to ATP and assumes a function in certain hormonal reactions
3 Cellular Organization
3.1 Cellular Organization
4 Tissue Organization
4.1 Introduction
4.1.1 Cells are highly organized units, but, in multicellular organisms, they do not function in isolation. They work together in a group of similar cells called a tissue.
4.2 Types of Tissues
4.2.1 A tissue is a group of similar cells and their intercellular substance that have a similar origin in an embryo and function together to perform a specialized activity.
4.2.2 The science that deals with the study of tissues is called histology (hiss’-TOL-ō-jē; histio=tissue; logos=study of). The various tissues of the body are classified into four principal types according to their function and structure:
4.2.2.1 Epithelial (ep'-i-T.HĒ-lē-al) tissue, which covers body surfaces, lines body cavities and ducts, and forms glands.
4.2.2.2 Connective tissue, which protects and supports the body and its organs, binds organs together, and stores energy reserves.
4.2.2.3 Muscular tissue, which is responsible for movement through the active generation of force.
4.2.2.4 Nervous tissue, which initiates, transmits, and interprets nerve impulses that coordinate body activities.
4.2.3 Epithelial tissue and connective tissue, except for bone and blood, will be discussed in detail in this chapter. The general features of bone tissue and blood will be introduced here, but their detailed discussion occurs later in the book. Similarly, the detailed discussion of muscle tissue and nervous tissue will be postponed.
4.3 Epithelial Tissue
4.3.1 Epithelial tissues perform many activities in the body, ranging from protection of underlying tissues against microbial invasion, drying out, and harmful environmental factors to secretion. Epithelial tissue, or more simply, epithelium, may be divided into two subtypes: (1) covering and lining epithelium and (2) glandular epithelium. Covering and lining epithelium forms the outer covering of external body surfaces and the outer covering of some internal organs. It lines body cavities and the interiors of the respiratory and gastrointestinal tracts, blood vessels, and ducts. It makes up, along with nervous tissue, the parts of the sense organs for smell, hearing, vision, and touch, which respond to stimuli. And it is the tissue from which gametes (sperm and eggs) develop. Glandular epithelium constitutes the secreting portion of glands.
4.3.2 Both types of epithelium consist largely or entirely of closely packed cells with little or no intercellular material between adjacent cells. (Such intercellular material is also called the matrix.) The points of attachment between adjacent plasma membranes of epithelial cells are called cell junctions. They not only provide cell-to-cell attachments but also inhibit the movement of materials into certain cells and provide channels for communication between other cells. Epithelial cells are arranged in continuous sheets that may be either single or multilayered. Nerves may extend through these sheets, but blood vessels do not. They are avascular (a = without; vascular = blood vessels). The vessels that supply nutrients and remove wastes are located in underlying connective tissue.
4.3.3 Both types of epithelium overlie and adhere firmly to the connective tissue, which holds the epithelium in position and prevents it from being torn. The attachment between the epithelium and the connective tissue is a thin extracellular layer called the basement membrane. With few exceptions, epithelial cells secrete along their basal surfaces a material consisting of a special type of collagen and glycoproteins. This structure averages 50 to 80 nm in thickness and is referred to as the basal lamina. Frequently, the basal lamina is reinforced by an underlying reticular lamina consisting of reticular fibers and glycoproteins. This lamina is produced by cells in the underlying connective tissue. The combination of the basal lamina and reticular lamina constitutes the basement membrane.
4.3.4 Within a tissue, most normal cells remain in place, anchored to basement membranes and connective tissues. In an adult, a few cells, such as phagocytes, routinely move through the intercellular material (matrix) during an infection. And, in an embryo, certain cells migrate extensively as part of the growth and development process. Various activities of cells depend on a variety of adhesion proteins found in the intercellular material and blood. These proteins interact with receptors on plasma membranes called integrins. Many adhesion proteins contain tripeptides (arginine, glycine, and aspartic acid) as the recognition site for integrins. Among the adhesion proteins are fibronectin, vitronectin, osteopontin, collagens, thrombospondin, and fibrinogen. Together, adhesion proteins and their integrins function in anchoring cells in position, providing traction for the movement of cells, differentiation, positioning of cells, and possibly growth.
4.3.5 Some tissues of the body are so highly differentiated (specialized) that they have lost their capacity for mitosis. Examples are muscle tissue and nervous tissue. Other tissues, such as epithelium, which are subjected to a certain amount of wear and tear and injury, have a continuous capacity for renewal because they contain stem cells that are capable of renewing the tissue. Examining cells that are sloughed off provides the basis for the Pap smear, a test for precancer and cancer diagnosis of the uterus, cervix, and vagina (Chapter 28).
4.3.6 Covering and Lining Epithelium
4.3.6.1 Arrangement of Layers
4.3.6.1.1 Covering and lining epithelium is arranged in several different ways related to location and function. If the epithelium is specialized for absorption or filtration and is in an area that has minimal wear and tear, the cells of the tissue are arranged in a single layer. Such an arrangement is called simple epithelium. If the epithelium is found, in an area with a high degree of wear and tear, then the / cells are stacked in several layers. This tissue is referred to as stratified epithelium. A third, less common arrangement of epithelium is called pseudostratified columnar. Like simple epithelium, pseudostratified epithelium has only one layer of cells. However, some of the cells do not reach the surface-an arrangement that gives the tissue a multilayered, or stratified, appearance. The cells in pseudostratified epithelium that do reach the surface either secrete mucus or contain cilia that move mucus and foreign particles for eventual elimination from the body.
4.3.6.2 Cell Shapes
4.3.6.2.1 In addition to classifying covering and lining epithelium according to the number of its layers, we may also categorize it by cell shape. The cells may be flat, cube like, columnar or a combination of shapes. Squamous_(SKWĀ-mus) cells are flattened and scale like. They are attached to each other and form a mosaic pattern. Cuboidal cells; are usually cube-shaped In cross section. They sometimes appear as hexagons. Columnar cells are tall and cylindrical, appearing as somewhat rectangular in shape when set on end. Transitional cells often have a combination of shapes and are found where there is a great degree of distention or expansion in the body. Transitional cells in the bottom layer of an epithelial tissue may range in shape from cuboidal to columnar. In the intermediate layer, they may be cuboidal or polyhedral. In the superficial layer, they may range from cuboidal to squamous, depending on how much they are pulled out of shape during certain body functions.
4.3.6.3 Classification
4.3.6.3.1 Considering layers and cell shapes in combination, we may classify covering and lining epithelillum as follows:
4.3.6.3.1.1 Simple
4.3.6.3.1.1.1 Squamous
4.3.6.3.1.1.2 Cuboidal
4.3.6.3.1.1.3 Columnar
4.3.6.3.1.2 Stratified
4.3.6.3.1.2.1 Squamous
4.3.6.3.1.2.2 Cuboidal
4.3.6.3.1.2.3 Columnar
4.3.6.3.1.2.4 Transitional
4.3.6.3.1.3 Pseudostratified columnar
4.3.6.3.1.3.1 Each of the epithelial tissues described in the following sections is illustrated in Exhibit 4-1
4.3.6.4 Simple Epithelium
4.3.6.4.1 Simple Squamous Epithelium
4.3.6.4.1.1 This type of simple epithelium consists of a single layer of flat, scale like cells. Its surface resembles a tiled floor. The nucleus of each cell is centrally located and oval or spherical. Since simple squamous epithelium has only one layer of cells, it is highly adapted to diffusion, osmosis, and filtration. Thus, it lines the air sacs of the lungs, where respiratory gases (oxygen and carbon dioxide) are exchanged between air spaces and blood. It is present in the part of the kidney that filters blood. It also lines the inner surfaces of the membranous labyrinth and tympanic membrane of the ear. Simple squamous epithelium is found in body parts that have little wear and tear.
4.3.6.4.1.2 Simple squamous epithelium that lines the heart, blood vessels, and lymph vessels and forms the walls of capillaries is known as endothelium. Simple squamous epithelium that forms the epithelial layer of serous membranes is called mesothelium. Serous membranes line the thoracic and abdominopelvic cavities and cover the viscera within them.
4.3.6.4.2 Simple Cuboidal Epithelium
4.3.6.4.2.1 Viewed from above, the cells of simple cuboidal epithelium appear as closely fitted polygons. The cuboidal nature of the cells is obvious only when the tissue is sectioned at right angles. Like simple squamous epithelium, these cells possess a central nucleus that is usually round. Simple cuboidal epithelium covers the surface of the ovaries, lines the anterior surface of the capsule of the lens of the eye, and forms the pigmented epithelium of the retina of the eye. In the kidneys, where it forms the kidney tubules and contains microvilli, it functions in water reabsorption. It also lines the smaller ducts of some glands and the secreting units of glands, such as the thyroid.
4.3.6.4.2.2 Simple cuboidal epithelium performs the functions of secretion and absorption. Secretion is the production and release by cells of a fluid that may contain a variety of substances such as mucus, perspiration, or enzymes. Absorption is the intake of fluids or other substances by cells of the skin or mucous membranes.
4.3.6.4.3 Simple Columnar Epithelium
4.3.6.4.3.1 The surface view of simple columnar epithelium is similar to that of simple cuboidal tissue. When sectioned at right angles to the surface, however, the cells appear somewhat rectangular. The nuclei, usually located near the bases of the cells, are commonly oval.
4.3.6.4.3.2 The luminal surfaces (surfaces adjacent to the lumen, or cavity of a hollow organ, vessel, or duct) of simple columnar epithelial cells are modified in several ways, depending on location and function. Simple columnar epithelium lines the gastrointestinal tract from the cardia of the stomach to the anus, the gallbladder, and excretory ducts of many glands. In such sites, the cells protect the underlying tissues. Many of the cells are also modified to
4.3.6.4.3.3
4.3.6.4.3.4 START
4.3.6.4.3.5 Exhibit 4-1 Epithelial Tissues
4.3.6.4.3.5.1
4.3.6.4.4 Stratified Cuboidal Epithelium
4.3.6.4.4.1
4.3.6.4.5 Stratified Columnar Epithelium
4.3.6.4.5.1
4.3.6.4.6 Transitional Epithelium
4.3.6.4.6.1
4.3.6.5 Pseudostratified Columnar Epithelium
4.3.6.6
4.3.7 Glandular Epithelium
4.3.7.1
4.3.7.2 Structural Classification of Exocrine Glands
4.3.7.2.1
4.3.7.3 Functional Classification of Exocrine Glands
4.3.7.3.1
4.4 Connective Tissue
4.4.1 Introduction
4.4.1.1 This is the most abundant tissue in the body and serves a binding and supporting function. With the exception of cartilage, which is Avascular, connective tissue is highly vascular with a rich blood supply. The cells of connective tissue are widely scattered as opposed to closely packed and there is considerable intercellular substance (matrix). In contrast to Epithelium, connective tissues do not occur on free surfaces, such as the surfaces of a body cavity or the external surface of the body. The general functions of connective tissues are protection, support, binding together various organs, separating structures such as skeletal muscles, and storage of reserve energy.
4.4.1.2 The tissue qualities are largely determined by the intercellular substance in a connective tissue. This tissue is a non-living substance and may consist of fluid, semi fluid, gel like, or fibrous material.
4.4.1.3 The intercellular material found in cartilage is firm but pliable. The intercellular material found in bone is considerably harder and not pliable.
4.4.1.4 Fibroblast (FĪ-brō-blast)
4.4.1.4.1 A large, flat cell that forms collagenous and elastic fibers and intercellular substance of loose connective tissue
4.4.1.5 The intercellular substances are produced by the cells of connective tissue (Fibroblasts).
4.4.1.6 These cells may also store fat, ingest bacteria and cell debris, form anticoagulants, or give rise to antibodies that protect against disease.
4.4.2 Classification
4.4.2.1.1 Connective tissues may be classified in several ways. We will classify them as follows:
4.4.2.1.2 Embryonic connective tissue
4.4.2.1.2.1 Mesenchvme
4.4.2.1.2.2 Mucous connective tissue
4.4.2.1.3
Adult connective
tissue
4.4.2.1.3.1 Connective tissue proper
4.4.2.1.3.1.1 Loose (areolar) connective tissue
4.4.2.1.3.1.2 Adipose tissue
4.4.2.1.3.1.3 Dense (collagenous) connective tissue
4.4.2.1.3.1.4 Elastic connective tissue
4.4.2.1.3.1.5 Reticular connective tissue
4.4.2.1.3.2 Cartilage
4.4.2.1.3.2.1 Hyaline cartilage
4.4.2.1.3.2.2
Fibrocartilage
4.4.2.1.3.2.3 Elastic cartilage
4.4.2.1.3.3 Osseous (bone) tissue
4.4.2.1.3.4 Vascular (blood) tissue
4.4.2.1.4 Each of the connective tissues described in the following sections is illustrated in Exhibit 4-2.
4.4.2.1.5
4.4.3 Embryonic Connective Tissue
4.4.3.1
Connective tissue that is present
primarily in the. embryo or fetus is called embryonic connective tissue. The
term embryo refers to a developing human from fertilization through the
first two months of pregnancy; a fetus refers to a developing human from
the third month of pregnancy to birth.
4.4.3.2 One example of embryonic connective tissue found almost exclusively in the embryo is mesenchyme (MEZen-kīm)-the tissue from which all other connective tissues eventually arise. Mesenchyme is located beneath the skin and along the developing bones of the embryo. Some mesenchymal cells are scattered irregularly throughout adult connective tissue, most frequently around blood vessels. Here mesenchymal cells differentiate into fibroblasts that assist in wound healing.
4.4.3.3 Another kind of embryonic connective tissue is mucous connective tissue (Wharton's jelly), found primarily in the fetus. This tissue is located in the umbilical cord of the fetus, where it supports the wall of the cord.
4.4.4 Adult Connective Tissue
4.4.4.1 Adult connective tissue is connective tissue that exists in the newborn that has differentiated from mesenchyme and that does not change after birth. It is subdivided into several kinds.
4.4.4.2 Exhibit 4-2 A
4.4.4.2.1 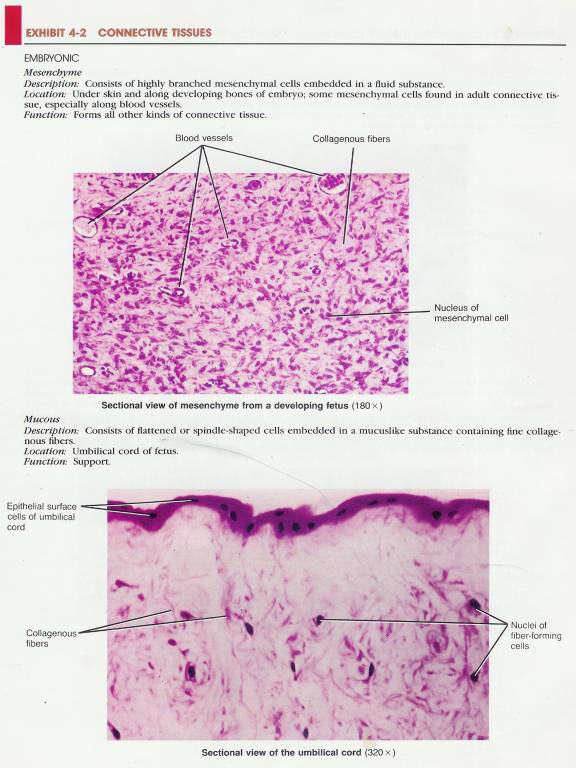
4.4.4.3 Exhibit 4-2 B
4.4.4.3.1 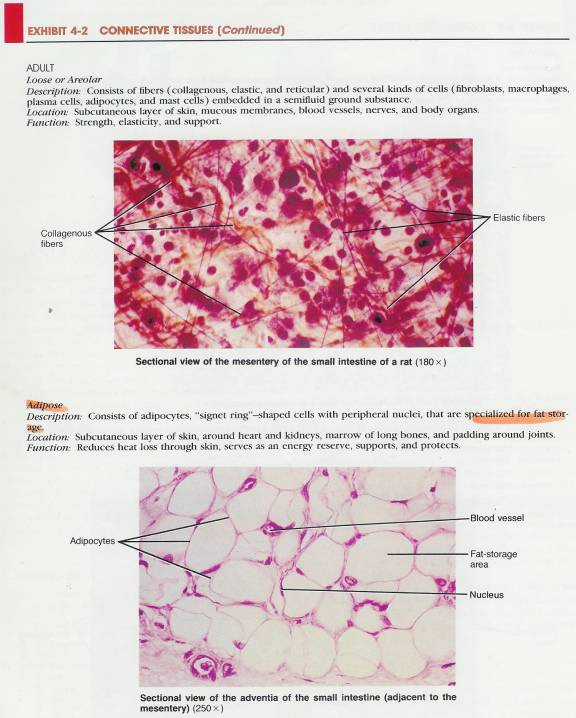
4.4.4.4 Exhibit 4-2 C
4.4.4.4.1 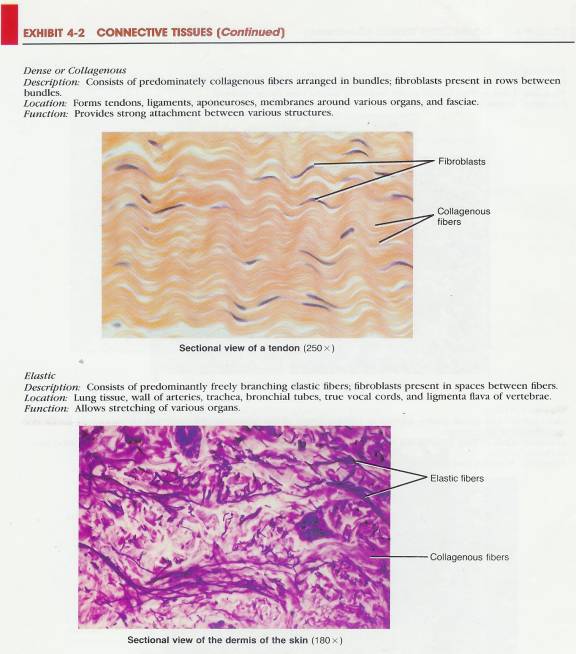
4.4.4.5 Exhibit 4-2 D
4.4.4.5.1 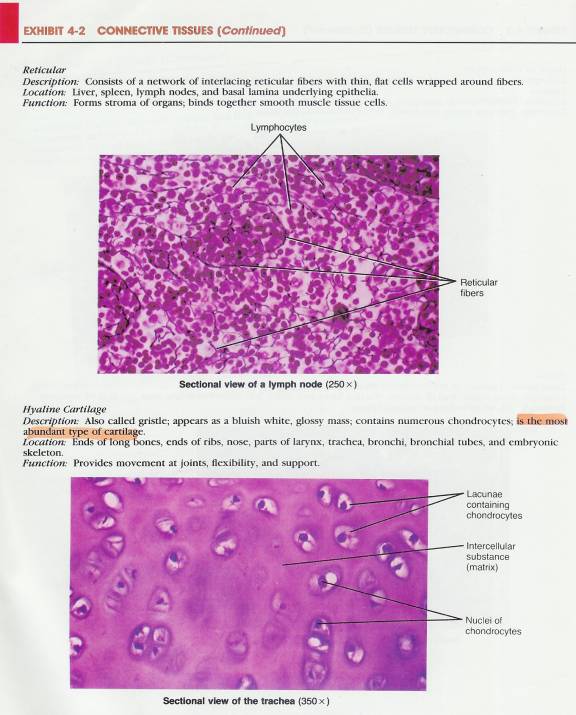
4.4.4.6 Exhibit 4-2 E
4.4.4.6.1 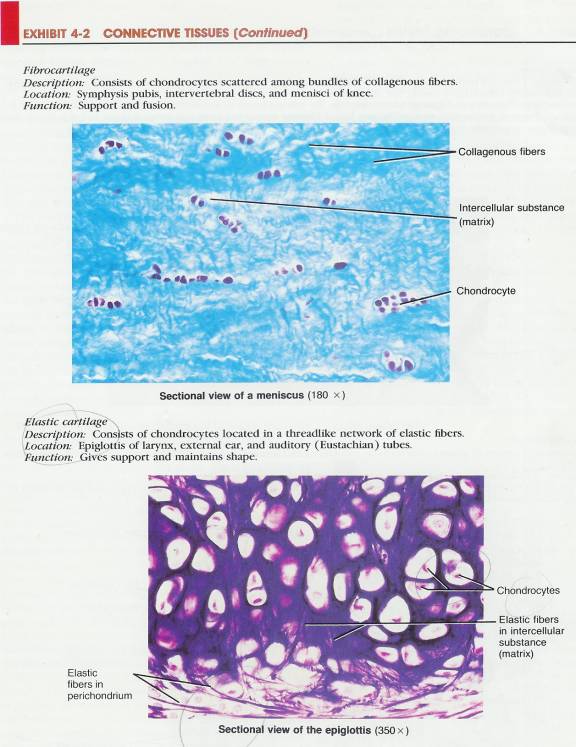
4.4.4.7 Exhibit 4-2 F
4.4.4.7.1 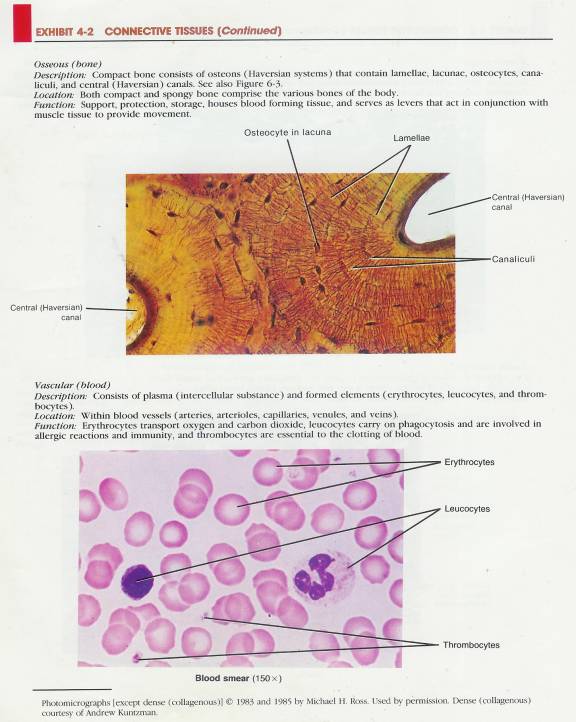
4.4.4.8 Connective Tissue Proper
4.4.4.8.1 There are five examples of connective tissue that has more or less fluid intercellular material and a fibroblast as the typical cell. All of this tissue is called connective tissue proper.
4.4.4.8.2 Loose (Areolar) Connective Tissue
4.4.4.8.2.1 Loose or areolar (a-RĒ-ō-Iar) connective tissue is one of the most widely distributed connective tissues in the body. Structurally, it consists of fibers and several kinds of cells embedded in a semifluid intercellular substance. The term loose refers to the loosely woven arrangement of fibers in the intercellular substance. The fibers are neither abundant nor arranged to prevent stretching.
4.4.4.8.2.2 The intercellular substance consists primarily of hyaluronic acid, chondroitin sulfate, dermatan sulfate, and keratan sulfate. It is secreted by connective tissue cells, mostly fibroblasts. The intercellular substance normally facilitates the passage of nutrients from the blood vessels of the connective tissue into adjacent cells and tissues, although the thick consistency of this acid may impede the movement of some drugs. But if an enzyme called hyaluronidase is injected into the tissue, the intercellular substance changes to a watery consistency. This feature is of clinical importance because the reduced viscosity hastens the absorption and diffusion of injected drugs and fluids through the tissue and thus can lessen tension and pain. Some bacteria, white blood cells, and sperm cells produce hyaluronidase.
4.4.4.8.2.3 The three types of
fibers embedded between the cells of loose connective tissue are collagenous,
elastic, and reticular fibers. Collagenous fibers are very tough and
resistant to a pulling force, yet allow some flexibility in the tissue because
!hey are not taut. These fibers often occur in bundles. They are composed of
many minute fibers called fibrils lying parallel to one another. The bundle
arrangement affords a great deal of strength. Chemically, collagenous fibers
consist of the protein collagen. Elastic fibers, by contrast, are
smaller than collagenous fibers and freely branch and rejoin one another.
Elastic fibers consist of a protein called elastin. These fibers also provide
strength and have great elasticity, up to 50 percent of their length. Reticular
fibers also consist of collagen, plus some glycoprotein. They are very thin
fibers that form branching networks. Like collagenous fibers, reticular fibers
provide support and strength and also form the stroma
(framework) of many soft organs.
4.4.4.8.2.4
The cells in loose connective tissue are numerous and varied.
Most are fibroblasts-large, flat cells with branching processes. If the
tissue is injured, fibroblasts are believed to form collagenous fibers, elastic
fibers, and the intercellular substance. When a fibroblast becomes relatively
inactive, it is sometimes referred to as a fibrocyte.
.
4.4.4.8.2.5 Other cells found in loose connective tissue are called fixed macrophages (MAK-r ō-fā-jēz; macro = large; phagein = to. eat), or histiocytes, which are derived from monocytes, a. type of white blood cell. Macrophages are irregular in form with short branching projections and are capable of engulfing bacteria and cellular debris by phagocytosis. Thus, they provide a vital defense for the body.
4.4.4.8.2.6 A third kind of cell in loose connective tissue is a plasma cell. It is small and either round or irregular and develops from a type of white blood cell called a lymphocyte (B cell). Plasma cells give rise to antibodies and, accordingly, provide a defense mechanism through immunity. They are found in many places in the body, but most are found in connective tissue, especially that of the gastrointestinal tract and the mammary glands.
4.4.4.8.2.7 Another cell in loose connective tissue is a mast cell. The mast cell is found in abundance along blood vessels. It forms heparin, an anticoagulant that prevents blood from clotting in the vessels. Mast cells are also believed to produce histamine and serotonin, chemicals that dilate small blood vessels.
4.4.4.8.2.8
Other cells in loose connective tissue include adipocytes
(fat cells) and leucocytes (white blood cells). .
4.4.4.8.2.9 Loose
connective tissue is continuous throughout the body. It is present in all
mucous membranes and around all blood vessels and nerves. And it occurs around
body organs and in the papillary (upper) region of the dermis of the skin.
Combined with adipose tissue, it forms the subcutaneous (sub'-kyoo-TĀ-nē-us;
sub = under; cut= skin) layer-the layer of tissue
that attaches the skin to underlying tissues and organs. The subcutaneous layer
is also referred to as the superficial fascia (FASH-ē-a) or hypodermis.
4.4.4.8.2.10
4.4.4.8.3 Clinical Application
4.4.4.8.3.1 
4.4.4.8.4 Adipose Tissue
4.4.4.8.4.1 Adipose Tissue is basically a form of loose connective tissue in which the cells, called adipocytes, are specialized for fat storage. Adipocytes are derived from fibroblasts, and the cells have the shape of a "signet ring" because the cytoplasm and nucleus are pushed to the edge of the cell by a large droplet of fat. Adipose tissue is found wherever loose connective tissue is located. Specifically, it is in the subcutaneous layer below the skin, around the kidneys, at the base and on the surface of the heart, in the marrow of long bones, as padding around joints, and behind the eyeball in the orbit. Adipose tissue is a poor conductor of heat and therefore reduces heat loss through the skin. It is a major energy reserve and generally supports and protects various organs.
4.4.4.8.4.2 Clinical Application
4.4.4.8.4.2.1 
4.4.4.8.5 Dense (Collagenous) Connective Tissue
4.4.4.8.5.1 Dense (Collagenous) Connective Tissue has fibers that are closely packed than found in loose connective tissue and the fibers can be irregularly or regularly arranged. These fibers are made up of collagen.
4.4.4.8.5.2 Collagen (KOL-a-jen
4.4.4.8.5.2.1 A protein that is the main organic constituent of connective tissue
4.4.4.8.5.3 Irregularly Arranged (fasciae)
4.4.4.8.5.3.1 When the tensions on the connective tissue are exerted in various directions, the fiber bundles are interwoven and without regular orientation. It forms most fasciae, the reticular (deeper) region of the dermis of the skin, the periosteum of bone, and the perichondrium of cartilage, and the membrane (fibrous) capsules around organs, such as the kidneys, liver, testes, and lymph nodes.
4.4.4.8.5.4 Regularly Arranged
4.4.4.8.5.4.1 This tissue is adapted for tension in one direction, and the fibers have an orderly, parallel arrangement.
4.4.4.8.5.4.2 The most common variety of dense regularly arranged connective tissue has a predominance of collagenous fibers arranged in bundles.
4.4.4.8.5.4.3 Fibroblasts are placed in rows between the bundles. The tissue is silvery white, tough, yet somewhat pliable. Because of its great strength, it is the principal component of tendons, which attach muscles to bones, aponeurosis (ap’-ō-noo-RŌ-sēz), which are sheetlike tendons connecting one muscle with another or with bone; and many ligaments (collagenous ligaments), which hold bones together at joints.
4.4.4.8.5.5 Tissue Example
4.4.4.8.5.5.1 Plantar Aponeurosis (PA) (A-paw-neurosis)
4.4.4.8.5.5.1.1 The plantar aponeurosis is Dense (Collagenous) Connective Tissue, which is regularly arranged and stretches across the sole of the foot similar to the palmar aponeurosis in the hand. It is a thickened layer of deep fascia, which serves both a protective and supportive function to the underlying muscles, vessels and nerves.
4.4.4.8.5.5.1.2 The fibers are oriented longitudinally and attached proximally to the calcaneus extending over its surface. It is attached distally to the metatarsophalangeal joints.
4.4.4.8.5.5.1.3 Functionally and histologically the plantar aponeurosis is tendon like and acts as an extension of the Achilles tendon. During the push off it is tightened along with the other plantar flexors, which pull on the Achilles tendon whose fibers may interdigitate with the PA. As push off progresses and the toes are extended the PA is further tightened. The tautness of the PA helps maintain the arch during running and walking activities. During stance phase the PA is less tight as intrinsic foot structures such as the long and short plantar ligament assume the burden of arch support.
4.4.4.8.5.5.1.4 The aponeurosis divides distally into five digital slips, one of which courses to each toe. Each division splits into two slips, which insert on either side of the plantar ligament of the MP joint
4.4.4.8.5.5.1.5 Fibers extend from the margins of the aponeurosis to cover partially both the medial and lateral plantar eminences.
4.4.4.8.5.5.1.6 The plantar aponeurosis and the long and short plantar ligaments are the main structures that support the longitudinal arch of the foot
4.4.4.8.5.5.2 Lateral Cord of the Plantar Aponeurosis
4.4.4.8.5.5.2.1 Helps support the lateral longitudinal arch. It is attached to the lateral Calcaneus blending with the plantar aponeurosis.
4.4.4.8.5.5.2.2 Distally this structure attaches to the base of the 5th metatarsal bone.
4.4.4.8.5.5.3 Plantar Fascia (fā-si-ă) (PF)
4.4.4.8.5.5.3.1 Extends over the medial and lateral side of the PA and covers the abductor Hallucis (hǎ-li-cis) on the medial side and abductor Digiti Minimi (dĭgĭtĭ mĭnĭmĭ) on the lateral side.
4.4.4.8.5.5.4 Long Plantar Ligament (ligamentum plantare longum; long calcaneocuboid ligament; superficial long plantar ligament) Illustration-Plantar Illustration-Medial
4.4.4.8.5.5.4.1 The rough surface in front of the calcaneus processes gives attachment to the long plantar ligament. On the distal end it attaches to the cuboid bone whose plantar surface presents in front with a deep groove (Peroneal Sulcus), which runs obliquely forward and medial ward (Peroneus Longus tendon inserts). Behind this groove is a prominent ridge where the long plantar ligament is attached.
4.4.4.8.5.5.4.2 The long plantar ligament is the longest of all the ligaments of the tarsus: it is attached behind to the plantar surface of the calcaneus in front of the tuberosity, and in front to the tuberosity on the plantar surface of the cuboid bone, the more superficial fibers being continued forward to the bases of the (sometimes second), third, fourth and fifth metatarsal bones. This ligament converts the groove on the plantar surface of the cuboid into a canal for the tendon of the Peroneus longus.
4.4.4.8.5.5.5 Plantar Calcaneocuboid Ligament (Short Plantar Ligament) (ligamentum calcaneocuboideum plantare; short calcaneocuboid ligament) Illustration-Plantar Illustration-Medial
4.4.4.8.5.5.5.1 The plantar calcaneocuboid ligament lies nearer to the bones than the preceding, from which it is separated by a little Areolar (ă-realer) tissue. It is a short but wide band of great strength, and extends from the tubercle and the depression in front of it, on the forepart of the plantar surface of the calcaneus, to the plantar surface of the cuboid behind the peroneal groove.
4.4.4.8.5.5.6 Plantar Calcaneonavicular (Spring) Ligament (ligamentum Calcaneonavicular plantare; inferior or internal calcaneonavicular ligament; calcaneonavicular ligament) Illustration-Plantar Illustration-Medial
4.4.4.8.5.5.6.1 The spring ligament passes from the anterior and medial margins of the sustentaculum tali forward to the inferior and inferomedial aspect of the Navicular. Its superior surface articulates with the underside of the head of the talus. This ligament maintains apposition of the medial aspects of the forefoot and Hindfoot and in so doing helps to maintain the normal arched configuration of the foot. Laxity of the ligament allows a medial separation between the calcaneus and forefoot, with the forefoot assuming an abducted position with respect to the Hindfoot.
4.4.4.8.5.5.6.2 At the same time, the foot is allowed to “untwist” which effectively lowers the normal arch of the foot, and the talar head is allowed to move medially and inferiorly.
4.4.4.8.5.5.6.3 The plantar calcaneonavicular ligament is a broad and thick band of fibers, which connects the anterior margin of the sustentaculum tali of the calcaneus to the plantar surface of the Navicular. This ligament not only serves to connect the calcaneus and Navicular, but supports the head of the talus, forming part of the articular cavity in which it is received. The dorsal surface of the ligament presents a fibrocartilaginous facet, lined by the synovial membrane, and upon this a portion of the head of the talus rests. Its plantar surface is supported by the tendon of the Tibialis posterior; its medial border is blended with the forepart of the deltoid ligament of the ankle-joint.
4.4.4.8.5.5.6.4 The plantar calcaneonavicular ligament, by supporting the head of the talus, is principally concerned in maintaining the arch of the foot. When it yields, the head of the talus is pressed downward, medialward, and forward by the weight of the body, and the foot becomes flattened, expanded, and turned lateralward, and exhibits the condition known as flat-foot. This ligament contains a considerable amount of elastic fibers, so as to give elasticity to the arch and spring to the foot; hence it is sometimes called the “spring” ligament. It is supported, on its plantar surface, by the tendon of the Tibialis posterior, which spreads out at its insertion into a number of fasciculi, to be attached to most of the tarsal and metatarsal bones. This prevents undue stretching of the ligament, and is a protection against the occurrence of flat-foot; hence muscular weakness is, in most cases, the primary cause of the deformity.
4.4.4.8.5.5.7 Retinaculum of foot/The Fascia Around the Ankle
4.4.4.8.5.5.7.1 Fibrous bands, or thickened portions of the fascia, bind down the tendons in front of and behind the ankle in their passage to the foot. They comprise three ligaments, viz., the transverse crural, the cruciate crural and the laciniate; and the superior and inferior peroneal Retinacula.
4.4.4.8.5.5.7.2 Transverse Crural Ligament (ligamentum transversum cruris; upper part of anterior annular ligament)
4.4.4.8.5.5.7.2.1 The transverse crural ligament binds down the tendons of Extensor digitorum longus, Extensor hallucis longus, Peroneus Tertius, and Tibialis anterior as they descend on the front of the tibia and fibula; under it are found also the anterior tibial vessels and deep peroneal nerve. It is attached laterally to the lower end of the fibula, and medially to the tibia; above it is continuous with the fascia of the leg.
4.4.4.8.5.5.7.3 Cruciate Crural Ligament (ligamentum cruciatum cruris; lower part of anterior annular ligament)
4.4.4.8.5.5.7.3.1 The cruciate crural ligament is a Y-shaped band placed in front of the ankle-joint, the stem of the Y being attached laterally to the upper surface of the calcaneus, in front of the depression for the interosseous talocalcaneal ligament; it is directed medialward as a double layer, one lamina passing in front of, and the other behind, the tendons of the Peroneus Tertius and Extensor digitorum longus. At the medial border of the latter tendon these two layers join together, forming a compartment in which the tendons are enclosed. From the medial extremity of this sheath the two limbs of the Y diverge: one is directed upward and medialward, to be attached to the tibial malleolus, passing over the Extensor hallucis longus and the vessels and nerves, but enclosing the Tibialis anterior by a splitting of its fibers. The other limb extends downward and medialward, to be attached to the border of the plantar aponeurosis, and passes over the tendons of the Extensor hallucis longus and Tibialis anterior and also the vessels and nerves.
4.4.4.8.5.5.7.4 Laciniate Ligament (ligamentum laciniatum; internal annular ligament)
4.4.4.8.5.5.7.4.1 The laciniate ligament is a strong fibrous band, extending from the tibial malleolus above to the margin of the calcaneus below, converting a series of bony grooves in this situation into canals for the passage of the tendons of the Flexor muscles and the posterior tibial vessels and tibial nerve into the sole of the foot. It is continuous by its upper border with the deep fascia of the leg, and by its lower border with the plantar aponeurosis and the fibers of origin of the Abductor hallucis muscle. Enumerated from the medial side, the four canals which it forms transmit the tendon of the Tibialis posterior; the tendon of the Flexor digitorum longus; the posterior tibial vessels and tibial nerve, which run through a broad space beneath the ligament; and lastly, in a canal formed partly by the talus, the tendon of the Flexor hallucis longus.
4.4.4.8.5.5.7.5 Peroneal Retinacula (Superior and Inferior)
4.4.4.8.5.5.7.5.1 The peroneal Retinacula are fibrous bands, which bind down the tendons of the Peronei longus and brevis as they run across the lateral side of the ankle. The fibers of the superior retinaculum (external annular ligament) are attached above to the lateral malleolus and below to the lateral surface of the calcaneus. The fibers of the inferior retinaculum are continuous in front with those of the cruciate crural ligament; behind they are attached to the lateral surface of the calcaneus; some of the fibers are fixed to the peroneal trochlea, forming a septum between the tendons of the Peronei longus and brevis.
4.4.4.8.5.5.7.6 Mucous Sheaths of the Tendons Around the Ankle
4.4.4.8.5.5.7.6.1 All the tendons crossing the ankle-joint are enclosed for part of their length in mucous sheaths, which have an almost uniform length of about 8 cm. each. On the front of the ankle (Fig. 441) the sheath for the Tibialis anterior extends from the upper margin of the transverse crural ligament to the interval between the diverging limbs of the cruciate ligament; those for the Extensor digitorum longus and Extensor hallucis longus reach upward to just above the level of the tips of the malleoli, the former being the higher. The sheath of the Extensor hallucis longus is prolonged on to the base of the first metatarsal bone, while that of the Extensor digitorum longus reaches only to the level of the base of the fifth metatarsal. On the medial side of the ankle (Fig. 442) the sheath for the Tibialis posterior extends highest up—to about 4 cm. above the tip of the malleolus—while below it stops just short of the tuberosity of the navicular. The sheath for Flexor hallucis longus reaches up to the level of the tip of the malleolus, while that for the Flexor digitorum longus is slightly higher; the former is continued to the base of the first metatarsal, but the latter stops opposite the first cuneiform bone. On the lateral side of the ankle (Fig. 441) a sheath which is single for the greater part of its extent encloses the Peronei longus and brevis. It extends upward for about 4 cm. above the tip of the malleolus and downward and forward for about the same distance.
4.4.4.8.6 Elastic Connective Tissue
4.4.4.8.6.1 Unlike collagenous connective tissue, elastic connective tissue has a predominance of freely branching elastic fibers. These fibers give the unstained tissue a yellowish color. Fibroblasts are present only in the spaces between the fibers. Elastic connective tissue can be stretched and will snap back into shape. It is a component of the walls of elastic arteries, trachea, bronchial tubes to the lungs, and the lungs themselves. Elastic connective tissue provides stretch and strength, allowing structures to perform their functions efficiently. Yellow elastic ligaments, as contrasted with collagenous ligaments, are composed mostly of elastic fibers; they form the ligamenta flava of the vertebrae (ligaments between successive vertebrae), the suspensory ligament of the penis, and the true vocal cords.
4.4.4.8.7 Reticular Connective Tissue
4.4.4.8.7.1 Reticular connective tissue consists of interlacing reticular fibers. It helps to form a delicate supporting stroma (framework) for many organs, including the liver, spleen, and lymph nodes. It is also found in the basal lamina underlying epithelia and around blood vessels and muscle. Reticular connective tissue also helps to bind together the fibers (cells) of smooth muscle tissue.
4.4.4.9 Cartilage
4.4.4.9.1 Cartilage is capable of enduring considerably more stress than the tissues just discussed. Unlike other connective tissues, cartilage has no blood vessels or nerves, except for those in the perichondrium (membranous covering). Cartilage consists of a dense network of collagenous fibers and elastic fibers firmly embedded in chondroitin sulfate, a jellylike intercellular substance (matrix). Whereas the strength of cartilage is due to its collagenous fibers, its resilience (ability to assume its original shape after deformation) is due to chondroitin sulfate. The cells of mature cartilage, called chondrocytes (KON-drō-sīts), occur singly or in groups within spaces called lacunae ( la-KOO-nē) in the intercellular substance. The surface of cartilage is surrounded by irregularly arranged dense. connective tissue called the perichondrium (per'-i-KON-drē-um; peri = around; chondro = cartilage). Three kinds of cartilage are recognized: hyaline cartilage, fibrocartilage, and elastic cartilage (Exhibit 4-2).
4.4.4.9.2 Hyaline Cartilage
4.4.4.9.2.1 This cartilage, also called gristle, 'appears in the body as a bluish-white, shiny substance. The collagenous fibers, although present, are not visible with ordinary staining techniques, and the prominent chondrocytes are found in lacunae. .Hyaline cartilage is the most abundant kind of cartilage in the body. It is found at joints over the ends of the long bones (where it is called articular cartilage) and forms the costal cartilages at the ventral ends of the ribs. Hyaline cartilage also helps to form the nose, larynx, trachea, bronchi, and bronchial tubes leading to the lungs. Most of the embryonic skeleton consists of hyaline cartilage. Hyaline cartilage affords flexibility and support and, as articular cartilage, reduces friction and absorbs shock.
4.4.4.9.3 Fibrocartilage
4.4.4.9.3.1 Chondrocytes scattered through many bundles of visible collagenous fibers are found in this type of cartilage. Fibrocartilage is found at the symphysis pubis, the point where the coxal (hip) bones fuse anteriorly at the midline. It is also found in the intervertebral discs between vertebrae and the menisci of the knee. This tissue combines strength and rigidity.
4.4.4.9.4 Elastic Cartilage
4.4.4.9.4.1 In this tissue, chondrocytes are located in a threadlike network of elastic fibers. Elastic cartilage provides strength and elasticity and maintains the shape of certain organs-the epiglottis of the larynx, the external part of the ear (pinna), and the auditory (Eustachian) tubes.
4.4.4.9.5 Growth of Cartilage
4.4.4.9.5.1 The growth of cartilage follows two basic patterns. In. interstitial (endogenous) growth, the cartilage increases rapidly in size through the division of existing chondrocytes and continuous deposition of increasing amounts of intercellular matrix by the chondrocytes. The formation of new chondrocytes and their production of new intercellular matrix causes the cartilage to expand from within-thus, the term interstitial growth. This growth pattern occurs while the cartilage is young and pliable-during childhood and adolescence.
4.4.4.9.5.2 In appositional (exogenous) growth, the growth of cartilage occurs because of the activity of the inner chondrogenic layer of the perichondrium. The deeper cells of the perichondrium, the fibroblasts, divide. Some differentiate into chondroblasts (immature cells that develop into specialized cells) and then into chondrocytes. As differentiation occurs, the chondroblasts become surrounded with intercellular matrix and become chondrocytes. As a result, the matrix is deposited on the surface of the cartilage, increasing its size. The new layer of cartilage is added beneath the perichondrium on the surface of the cartilage, causing it to grow in width. Appositional growth starts later than interstitial growth and continues throughout life.
4.4.4.10 Osseous Tissue (Bone)
4.4.4.10.1 Together, cartilage, joints, and osseous (OS-ē-us) tissue (bone) comprise the skeletal system. Mature bone cells are called osteocytes. The intercellular substance consists of mineral salts, primarily calcium phosphate and calcium carbonate, and collagenous fibers. The salts are responsible for the hardness of bone.
4.4.4.10.2
Bone tissue is classified as either compact (dense) or
spongy (cancellous), depending on how the intercellular substance and cells are
organized. At this point, we will discuss compact bone only. The basic unit of
compact bone is called an osteon
(Haversian system). Each osteon consists of lamellae, concentric rings of hard,
intercellular substance; lacunae,
small spaces between lamellae that contain osteocytes; canaliculi, radiating minute canals
that provide numerous routes so that nutrients can reach osteocytes and wastes
can be removed from them; and a central
(Haversian) canal that contains blood vessels and nerves. Notice that
whereas osseous tissue is vascular, cartilage is not. Also, the lacunae of
osseous tissue are interconnected by canaliculi; those of cartilage are not.
4.4.4.10.3
Functionally, the skeletal system supports soft
tissues, protects delicate structures, works with skeletal muscles to
facilitate movement, stores calcium and phosphorus, houses red marrow, which
produces several kinds of blood cells, and houses yellow marrow, which contains
lipids as an energy source.
4.4.4.10.4 The details of compact and spongy bone are discussed in Chapter 6.
4.4.4.11 Vascular Tissue (Blood)
4.4.4.11.1 Vascular tissue (blood) is a liquid connective tissue that consists of an intercellular substance called plasma and formed elements (cells and cell-like structures).
4.4.4.11.2 Plasma is a straw-colored liquid that consists mostly of water plus some dissolved substances (nutrients, enzymes, hormones, respiratory gases, and ions). The formed elements are erythrocytes (red blood cells), leucocytes (white blood cells), and thrombocytes (platelets). The fibers characteristic of all connective tissues are present in blood only when it is clotted.
4.4.4.11.3 Erythrocytes function in transporting oxygen to body cells and removing carbon dioxide from them. Leucocytes are involved in phagocytosis, immunity, and allergic reactions. Thrombocytes function in blood clotting.
4.4.4.11.4 The details of blood are considered in Chapter 19.
4.5 Membranes
4.5.1
The combination of an epithelial layer
and an underlying connective tissue layer constitutes an epithelial membrane. The
principal, epithelial membranes of the body are mucous membranes, serous
membranes, and the cutaneous membrane or skin.
Another kind of membrane, a synovial membrane, does not contain epithelium.
4.5.2 Mucous Membranes
4.5.2.1 A mucous membrane, or
mucosa, lines a body cavity that opens
directly to the exterior. Mucous membranes line the entire gastrointestinal,
respiratory, excretory, and reproductive tracts (see Figure.24- 2) and consist
of a lining layer of epithelium and an underlying layer of connective tissue.
In addition, most mucous membranes also contain a layer of smooth muscle called
the muscularis mucosae.
4.5.2.2 The epithelial layer of a mucous membrane secretes mucus, which prevents the cavities from drying out. It also traps particles in the respiratory passageways and lubricates food as it moves through the gastrointestinal tract. In addition, the epithelial layer is responsible for the secretion of digestive enzymes and the absorption of food.
4.5.2.3 The connective tissue layer of a mucous membrane is called the lamina propria. The lamina propria is so named because it belongs to the mucous membrane (proprius = one's own). It binds the epithelium to the underlying structures and allows some flexibility of the membrane. It also holds the blood vessels in place, protects underlying muscles from abrasion or puncture, and allows the diffusion of oxygen and nutrients to the epithelium covering it, and the diffusion of carbon dioxide and wastes from the epithelium covering it.
4.5.2.4 The muscularis mucosae contains smooth muscle fibers and separates the mucosa from the submucosa beneath. Its role in the gastrointestinal tract is considered in Chapter 24.
4.5.3 Serous Membranes
4.5.3.1 A serous membrane, or
serosa, lines a body cavity that does not
open directly to the exterior, and it covers the organs that lie within the
cavity. Serous membranes consist of thin layers of loose connective tissue
covered by a layer of mesothelium, and they are composed of two portions. The
part attached to the cavity wall is called the parietal (pa-RĪ-e-tal) portion. The part that covers and
attaches to the organs inside these cavities is the visceral portion. The serous
membrane lining the thoracic cavity and covering the lungs is called the pleura (see Figure 1-7). The
membrane lining the heart cavity and covering the heart is the pericardium (cardio = heart): The serous membrane lining the abdominal cavity and covering
the abdominal organs and some pelvic organs is called the peritoneum.
4.5.3.2 The epithelial layer of a serous membrane secretes a lubricating fluid, called serous fluid, that allows the organs to glide easily against one another or against the walls of the cavities. The connective tissue layer of the serous membrane consists of a relatively thin layer of loose connective tissue.
4.5.4 Cutaneous Membrane
4.5.4.1 The cutaneous membrane, or skin, constitutes an organ of the integumentary system and is discussed in the next chapter.
4.5.5 Synovial Membranes
4.5.5.1 Synovial membranes line the cavities of the freely movable joints (see Figure 9-1a). Like serous membranes, they line structures that do not open to the exterior. Unlike mucous, serous, and cutaneous membranes, they do not contain epithelium and are therefore not epithelial membranes. They are composed of loose connective tissue with elastic fibers and varying amounts of fat. Synovial membranes secrete synovial fluid, which lubricates the articular cartilage at the ends of bones as they move at joints and nourishes the articular cartilage covering the bones that form the joints. These are articular synovial membranes. Other synovial membranes line cushioning sacs, called bursae, and tendon sheaths in our hands and feet that facilitate the movement of muscle tendons.
4.6 Muscle Tissue
4.6.1 Muscle tissue consists of fibers (cells) that are highly specialized for the active generation of force for contraction. As a result of this characteristic, muscle tissue provides motion, maintenance of posture, and heat production. On the basis of certain structural and functional characteristics, muscle tissue is classified into three types: skeletal, cardiac, and smooth (Exhibit 4-3).
4.6.2 Skeletal muscle tissue is named for its location attached to bones. It is also striated; that is, the fibers (cells) contain alternating light and dark bands (striations) that are perpendicular to the long axes of the fibers. The striations are visible under a microscope. Skeletal muscle tissue is also voluntary because it can be made to contract or relax by conscious control. A single skeletal muscle fiber is cylindrical, and the fibers are parallel to each other in a tissue. Each muscle fiber contains a plasma membrane, the sarcolemma, surrounding the cytoplasm, or sarcoplasm. Skeletal muscle fibers are multinucleate (they have more than one nucleus), and the nuclei lie close to the sarcolemma. The contractile elements of skeletal muscle fibers are proteins called myofilaments. They contain wide, transverse, dark bands and narrow light ones that give the fibers the striated appearance.
4.6.3 Cardiac muscle tissue forms the bulk of the wall of the heart. Like skeletal muscle
tissue, it is striated. However, unlike skeletal muscle tissue, it is usually
involuntary; its contraction is usually not under conscious control. Cardiac
muscle fibers are roughly quadrangular and branch to form networks throughout
the tissue. The fibers usually have only one nucleus that is centrally located.
Cardiac muscle fibers are bound to each other by transverse thickenings of the
sarcolemma called intercalated discs. These are unique to cardiac muscle and serve to strengthen the
tissue and aid muscle action potential conduction by way of channels called gap junctions.
4.6.4 Smooth muscle tissue is located in the walls of hollow internal structures such as blood vessels, the stomach, intestines, and urinary bladder. Smooth muscle fibers are usually involuntary, and they are nonstriated (smooth). Each smooth muscle fiber is thickest in the midregion, with either end tapering to a point, and contains a single, centrally located nucleus.
4.6.5 A more detailed discussion of muscle tissue is considered in Chapter 10.
4.6.6 Exhibit 4-3
4.6.6.1
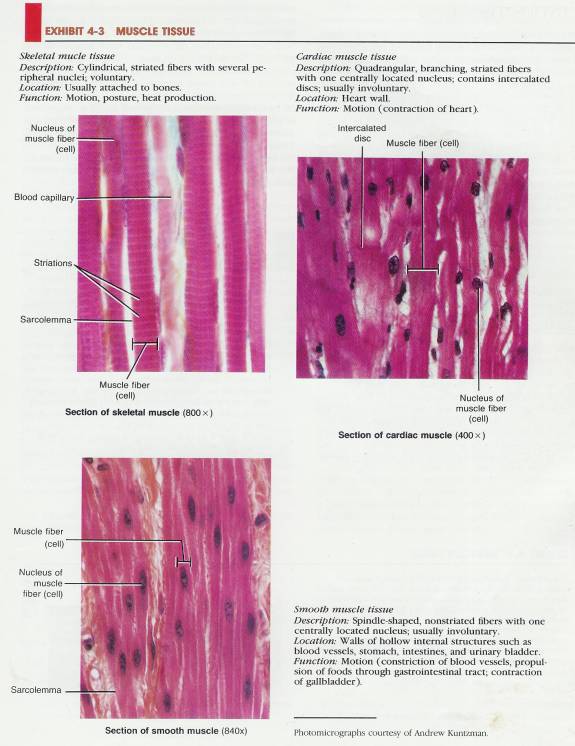
4.7 Nervous Tissue
4.7.1 Despite the tremendous complexity of the nervous system, it consists of only two principal kinds of cells: neurons and neuroglia. Neurons, or nerve cells, are highly specialized cells that are sensitive to various stimuli; converting stimuli to nerve impulses; and conducting nerve impulses to other neurons, muscle fibers, or glands. Neurons are the structural and functional units of the nervous system. Most consist of three basic portions: cell body and two kinds of processes called dendrites and axons (Exhibit 4-4). The cell body (perikaryon) contains the nucleus and other organelles. Dendrites are highly branched processes of the cell body that conduct nerve impulses toward the cell body. Axons are single, long processes of the cell body that conduct nerve impulses away from the cell body.
4.7.2 Neuroglia are cells that protect and support neurons. They are of clinical interest because they are frequently the sites of tumors of the nervous system.
4.7.3 The detailed structure and function of neurons and neuroglia are considered in Chapter 12.
4.8 Tissue Repair: An Attempt to Restore Homeostasis
4.8.1 Tissue repair is the process by which new tissues replace dead or damaged cells. New cells originate by cell duplication from the stroma, the supporting connective tissue, or from the parenchyma, cells that form the organ's functioning part. The epithelial cells that secrete and absorb are the parenchymal cells of the intestine, for example. The restoration of an injured organ or tissue to normal structure and function depends entirely on which type of cell-parenchymal or stromal-is active in the repair. If only parenchymal elements accomplish the repair, a perfect or near -perfect reconstruction of the injured tissue may occur. However, if the fibroblast cells of the stroma are active in the repair, the tissue will be replaced with new connective tissue called scar tissue. In this process, fibroblasts synthesize collagen and protein polysaccharides that aggregate to form scar tissue. The process of scar tissue formation is known as fibrosis. Since scar tissue is not specialized to perform the functions of the parenchymal tissue, the overall function of the tissue is impaired. If the rate of collagen breakdown in a scar exceeds production, the scar becomes softer and less bulky. If, on the other hand, the rate of collagen production exceeds breakdown, a keloid (hypertrophic) scar develops. Such a scar is sharply elevated, irregularly shaped, and unsightly.
4.8.2 Clinical Application
4.8.2.1

4.8.3 Exhibit 4-4
4.8.3.1
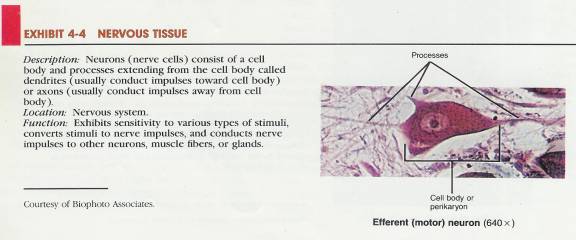
4.8.4 The cardinal factor in tissue repair-lies in the capacity of parenchymal tissue to regenerate. This capacity, in turn, depends on the ability of the parenchymal cells to replicate quickly.
4.8.5 Repair Process
4.8.5.1 If injury to a tissue is slight, repair may sometimes be
accomplished with the drainage and reabsorption of pus (accumulation of
leucocytes and fluid resulting from inflammation), followed by parenchymal
regeneration. When the area of skin loss is great, fluid moves out of the
capillaries and the area becomes dry. Fibrin, an insoluble protein in a blood
clot, seals the open tissue by hardening into a scab.
4.8.5.2 When tissue and cell damage are extensive and severe, as in large, open wounds, both the connective tissue stroma and the parenchymal cells are active in repair. This repair involves the rapid cell division of many fibroblasts, the manufacture of new collagenous fibers to provide strength, and an increase by cell division of the number of small blood vessels in the area. All these processes create an actively growing connective tissue called granulation tissue. This new granulation tissue forms across a wound or surgical incision to provide a framework (stroma). The framework supports the epithelial cells that migrate into the open area and fill it. The newly formed granulation tissue also secretes a fluid that kills bacteria.
4.8.6 Conditions Affecting Repair
4.8.6.1 Three factors affect tissue repair: nutrition, blood circulation, and age. Nutrition is vital in the healing process since a great demand is placed on the body's store of nutrients. Protein-rich diets are important since most of the cell structure is made from proteins. Vitamins also play a direct role in wound healing. Among the vitamins involved and their roles in wound healing are the following:
4.8.6.1.1
Vitamin A is essential in the replacement of
epithelial tissues, especially in the respiratory tract.
4.8.6.1.2 The B vitamins-thiamine, nicotinic acid, riboflavin-are coenzymes needed by many enzyme systems in cells. They are needed especially for the enzymes involved in decomposing glucose to CO2 and H2O, which is crucial to both heart and nervous tissue. These vitamins may relieve pain in some cases and are necessary for division of the cells that accomplish repair.
4.8.6.1.3 Vitamin C directly affects the normal production and maintenance of intercellular substances. It is required for the manufacture of cementing elements of connective tissues, especially collagen. Vitamin C also strengthens and promotes the formation of new blood vessels. With vitamin C deficiency, even superficial wounds fail to heal, and the walls of the blood vessels become fragile and are easily ruptured.
4.8.6.1.4 Vitamin D is necessary for the proper absorption of calcium from the intestine. Calcium gives bones their hardness and is necessary for the healing of fractures.
4.8.6.1.5 Vitamin E is believed to promote healing of injured tissues and may prevent scarring.
4.8.6.1.6 Vitamin K assists in the clotting of blood and thus prevents the injured person from bleeding to death. Although vitamin K is essential for the formation of certain clotting factors, it is not, itself, a coagulant. The body, with the aid of colon bacteria, normally synthesizes its own vitamin K. In humans, bacteria in the colon synthesize vitamin K which can be' absorbed and utilized.
4.8.6.2
In
tissue repair, proper blood circulation is indispensable. It is the blood that
transports oxygen, nutrients, antibodies, and many defensive cells to the site
of injury. The blood also plays an important role in the removal of tissue
fluid, blood cells that have been depleted of oxygen, bacteria, foreign bodies,
and debris. These elements would otherwise interfere with healing.
4.8.6.3
Generally,
tissues heal faster and leave less obvious scars in the young than in the aged.
The young body is generally in a much better nutritional state, its tissues
have a better blood supply, and the cells of younger people have a faster
metabolic rate. Thus, cells can duplicate their materials and divide more
quickly.
4.8.7 Summary
4.8.7.1 Types of Tissues
4.8.7.1.1 1. A tissue is a group of similar cells and their intercellular substance that have a similar embryological origin and are specialized for a particular function.
4.8.7.1.2 2. Depending on their function and structure, the various tissues of the body are classified into four principal types: epithelial, connective, muscular, and nervous.
4.8.7.2 Epithelial Tissue
4.8.7.2.1 1. Epithelium has many cells, little intercellular material, and no blood vessels (avascular). It is attached to connective tissue by a basement membrane. It can replace itself.
4.8.7.2.2 2. The subtypes of epithelium include covering and lining epithelium and glandular epithelium.
4.8.7.3 Covering and Lining Epithelium (p. 93)
4.8.7.3.1 1. Layers are arranged as simple (one layer), stratified (several layers), and pseudostratified (one layer that appears as several); cell shapes include squamous (flat), cuboidal (cubelike), columnar (rectangular), and transitional (variable).
4.8.7.3.2 2. Simple squamous epithelium is adapted for diffusion and filtration and is found in lungs and kidneys. Endothelium lines the heart and blood vessels. Mesothelium lines the thoracic and abdominopelvic cavities and covers the organs within' them.
4.8.7.3.3 3. Simple cuboidal epithelium is adapted for secretion and absorption. It is found covering ovaries, in kidneys and eyes, and lining some glandular ducts.
4.8.7.3.4 4. Nonciliated simple columnar epithelium lines most of the gastrointestinal tract. Specialized cells containing microvilli perform absorption. Goblet cells secrete mucus. In a few portions of the respiratory tract, the cells are ciliated to move foreign particles trapped in mucus out of the body.
4.8.7.3.5 5. Stratified squamous epithelium is protective. It lines the upper gastrointestinal tract and vagina and forms the outer layer of skin.
4.8.7.3.6 Stratified cuboidal epithelium is found in adult sweat glands, pharynx, epiglottis, and portions of the urethra.
4.8.7.3.7 7. Stratified columnar epithelium protects and secretes. It is found in the male urethra and large excretory ducts.
4.8.7.3.8 8. Transitional epithelium lines the urinary bladder and is capable of stretching.
4.8.7.3.9 9. Pseudostratified columnar epithelium has only one layer but gives the appearance of many. It lines larger excretory ducts, parts of urethra, auditory (Eustachian) tubes, and most upper respiratory structures, where it protects and secretes.
4.8.7.4 Glandular Epithelium (p. 100)
4.8.7.4.1 A gland is a single cell or a mass of epithelial cells adapted for secretion.
4.8.7.4.2 Exocrine glands (sweat, oil, and digestive glands) secrete into ducts or directly onto a free surface.
4.8.7.4.3 Structural classification includes unicellular and multicellular glands; multicellular glands are further classified as tubular, acinar, tubuloacinar, simple, and compound.
4.8.7.4.4 Functional classification includes holocrine, merocrine, and apocrine glands.
4.8.7.4.5 Endocrine glands secrete hormones into the blood.
4.8.7.5 Connective Tissue (p. 102)
4.8.7.5.1 Connective tissue is the most abundant body tissue. It has few cells, an extensive intercellular substance, and a rich blood supply (vascular), except for cartilage. It does not occur on free surfaces.
4.8.7.5.2 2. The intercellular substance determines the tissue's qualities.
4.8.7.5.3 Connective tissue protects, supports, and binds organs together.
4.8.7.5.4 4. Connective tissue is classified into two principal types: embryonic and adult.
4.8.7.5.5 Embryonic Connective Tissue (p. 102) .
4.8.7.5.5.1 Mesenchyme forms all other connective tissues.
4.8.7.5.5.2 Mucous connective tissue is found in the umbilical cord of the fetus, where it gives support.
4.8.7.5.6 Adult Connective Tissue (p. 102)
4.8.7.5.6.1 Adult connective tissue is connective tissue that differentiates from mesenchyme and exists in the newborn and does not change after birth. It is subdivided into several kinds: connective tissue proper, cartilage, bone tissue, and vascular tissue.
4.8.7.5.6.2 Connective tissue proper has a more or less fluid intercellular material, and a typical cell is the fibroblast. Five examples of such tissues may be distinguished.
4.8.7.5.6.3 Loose (areolar) connective tissue is one of the most widely distributed connective tissues in the body. Its intercellular substance (hyaluronic acid) contains fibers (collagenous, elastic, and reticular) and various cells (fibroblasts, macrophages, plasma, and mast). Loose connective tissue is found in all mucous membranes, around body organs, and in the subcutaneous layer.
4.8.7.5.6.4 Adipose tissue is a form of loose connective tissue in which the cells, called adipocytes, are specialized for fat storage. It is found in the subcutaneous layer and around various organs.
4.8.7.5.6.5 Dense (collagenous) connective tissue has a close packing of fibers (regularly or irregularly arranged). It is found as a component of fascia, membranes of organs, tendons, ligaments, and aponeuroses.
4.8.7.5.6.6 Elastic connective tissue has a predominance of freely branching elastic fibers that give it a yellow color. It is found in elastic arteries, trachea, bronchial tubes, and true vocal cords.
4.8.7.5.6.7 Reticular connective tissue consists of interlacing reticular fibers and forms the stroma of the liver, spleen, and lymph nodes.
4.8.7.5.6.8
8. Cartilage has a jellylike matrix containing
collagenous and " elastic fibers and chondrocytes.
4.8.7.5.6.9 Hyaline cartilage is found in the embryonic skeleton, at the ends of bones, in the nose, and in respiratory structures. It is flexible, allows movement, and provides support.
4.8.7.5.6.10 Fibrocartilage connects the pelvic bones and the vertebrae. It provides strength.
4.8.7.5.6.11 Elastic cartilage maintains the shape of organs such as the larynx, auditory (Eustachian) tubes, and external ear.
4.8.7.5.6.12 The growth of cartilage is accomplished by interstitial growth (from within) and appositional growth (from without).
4.8.7.5.6.13 Osseous tissue (bone) consists of mineral salts and collagenous fibers that contribute to the hardness of bone and cells called osteocytes. It supports, protects, helps provide movement, stores minerals, and houses red marrow.
4.8.7.5.6.14 Vascular tissue (blood) consists of plasma and formed elements (erythrocytes, leucocytes, and thrombocytes). Functionally, its cells transport, carry on phagocytosis, participate in allergic reactions, provide immunity, and bring about blood clotting.
4.8.7.6 Membranes (p.111)
4.8.7.6.1 1. An epithelial membrane is an epithelial layer overlying a connective tissue layer. Examples are mucous, serous, and cutaneous membranes.
4.8.7.6.2 2. Mucous membranes line cavities that open to the exterior, such as the gastrointestinal tract.
4.8.7.6.3 Serous membranes (pleura, pericardium, peritoneum) line closed cavities and cover the organs in the cavities. These membranes consist of parietal and visceral portions.
4.8.7.6.4 The cutaneous membrane is the skin.
4.8.7.6.5 Synovial membranes line joint cavities, bursae, and tendon sheaths and do not contain epithelium.
4.8.7.7 Muscle Tissue (p. 112)
4.8.7.7.1 1. Muscle tissue is modified for contraction and thus provides motion, maintenance of posture, and heat production,
4.8.7.7.2 2, Skeletal muscle tissue is attached to bones, is striated, and is voluntary,
4.8.7.7.3 3. Cardiac muscle tissue forms most of the heart wall, is striated, and is usually involuntary.
4.8.7.7.4 Smooth muscle tissue is found in the walls of hollow internal structures (blood vessels and viscera), is nonstriated, and is usually involuntary.
4.8.7.8 Nervous Tissue (p. 114)
4.8.7.8.1 The nervous system is composed of neurons (nerve cells) and neuroglia (protective and supporting cells).
4.8.7.8.2 2. Most neurons consist of a cell body and two types of processes called dendrites and axons.
4.8.7.8.3 3. Neurons are sensitive to stimuli, convert stimuli into nerve impulses, and conduct nerve impulses.
4.8.7.9
Tissue Repair: An Attempt to Restore Homeostasis (p. 114)
4.8.7.9.1 1. Tissue repair is the replacement of damaged or destroyed cells by healthy ones.
4.8.7.9.2 It begins during the active phase of inflammation and is not completed until after harmful substances in the inflamed area have been neutralized or removed.
4.8.7.9.3 Repair Process (p. 114)
4.8.7.9.3.1 If the injury is superficial, tissue repair involves pus removal (if pus is present), scab formation, and parenchymal regeneration.
4.8.7.9.3.2 If damage is extensive, granulation tissue is involved.
4.8.7.9.4 Conditions Affecting Repair (p. 115)
4.8.7.9.4.1 Nutrition is important to tissue repair. Various vitamins (A, some B, D, C, E, and K) and a protein-rich diet are needed.
4.8.7.9.4.2 Adequate circulation of blood is needed.
4.8.7.9.4.3 The tissues of young people repair rapidly and efficiently; the process slows down with aging.
5 Integumentary System*
5.1
Skin
5.1.1
Physiology
5.1.2
Structure
5.1.2.1
Epidermis
5.1.2.1.1
Stratum
Basale
5.1.2.1.2
Stratum
Spinosum
5.1.2.1.3
Stratum Granulosum
5.1.2.1.4
Stratum
Lucidum
5.1.2.1.5
Stratum
Comeum
5.1.2.2
Dermis
5.1.2.3
Skin
Color
5.1.2.4
Epidermal
Ridges and Grooves
5.1.3
Skin
Wound Healing: Restoration of Homeostasis
5.1.3.1
Epidermal
Wound Healing
5.1.3.2
Deep
Wound Healing
5.1.4
Epidermal
Derivatives
5.1.4.1
Hair
5.1.4.2
Glands
5.1.4.2.1
Sebaceous
(Oil) Glands
5.1.4.2.2
Sudoriferous (Sweat)
Glands
5.1.4.2.3
Ceruminous
Glands
5.1.5
Homeostasis
of Body Temperature
5.1.6
Aging
and The Integumentary System
5.1.7
Developmental
Anatomy of the Integumentary System
6 Skeletal Tissue
7
Skeletal
System-Axial*

7.1
Axial
Skeleton
7.1.1
Types
of Bones
7.1.2
Surface Markings
Back Table
of Contents References
7.1.2.1
Introduction
7.1.2.1.1
7.1.2.2
Depressions
and Openings
7.1.2.2.1
Fissure
(FISH-ur)
7.1.2.2.1.1 Description
7.1.2.2.1.1.1
7.1.2.2.1.2 Example
7.1.2.2.1.2.1
7.1.2.2.2
Foramen
(fō-RĀ-men; foramen=hole)
7.1.2.2.2.1 Description
7.1.2.2.2.1.1
7.1.2.2.2.2 Example
7.1.2.2.2.2.1
7.1.2.2.3
Meatus
(mē-Ā-tus; meatus=canal)
7.1.2.2.3.1 Description
7.1.2.2.3.1.1
7.1.2.2.3.2 Example
7.1.2.2.3.2.1
7.1.2.2.4
Paranasal
Sinus (sin=cavity)
7.1.2.2.4.1 Description
7.1.2.2.4.1.1
7.1.2.2.4.2 Example
7.1.2.2.4.2.1
7.1.2.2.5
Groove or
Sulcus (Sulcus=ditchlike groove)
7.1.2.2.5.1 Description
7.1.2.2.5.1.1
7.1.2.2.5.2 Example
7.1.2.2.5.2.1
7.1.2.2.6
Fossa
(fossa=basinlike depression)
7.1.2.2.6.1 Description
7.1.2.2.6.1.1
7.1.2.2.6.2 Example
7.1.2.2.6.2.1
7.1.2.3
Processes
Back Table of Contents References
7.1.2.3.1
Condyle
(KON-dīl; condylus=knucklelike process)
7.1.2.3.1.1 Description
7.1.2.3.1.1.1
7.1.2.3.1.2 Example
7.1.2.3.1.2.1
7.1.2.3.2
Head
7.1.2.3.2.1 Description
7.1.2.3.2.1.1
7.1.2.3.2.2 Example
7.1.2.3.2.2.1
7.1.2.3.3
Facet
7.1.2.3.3.1 Description
7.1.2.3.3.1.1
7.1.2.3.3.2 Example
7.1.2.3.3.2.1
7.1.2.3.4
Tubercle
(TOO-ber-kul; tube=knob)
7.1.2.3.4.1 Description
7.1.2.3.4.1.1
7.1.2.3.4.2 Example
7.1.2.3.4.2.1
7.1.2.3.5 Tuberosity
7.1.2.3.5.1 Description
7.1.2.3.5.1.1 A large, rounded, usually roughened process
7.1.2.3.5.2 Example
7.1.2.3.5.2.1 Ischial tuberosity of the coxal (hip) bone
7.1.2.3.6
Trochanter
(trō-KAN-ter)
7.1.2.3.6.1 Description
7.1.2.3.6.1.1
7.1.2.3.6.2 Example
7.1.2.3.6.2.1
7.1.2.3.7
Line
7.1.2.3.7.1 Description
7.1.2.3.7.1.1
7.1.2.3.7.2 Example
7.1.2.3.7.2.1
7.1.2.3.8
Spinous
Process (spine)
7.1.2.3.8.1 Description
7.1.2.3.8.1.1
7.1.2.3.8.2 Example
7.1.2.3.8.2.1
7.1.2.3.9
Epicondyle
(epi=above)
7.1.2.3.9.1 Description
7.1.2.3.9.1.1
7.1.2.3.9.2 Example
7.1.2.3.9.2.1
7.1.3
Divisions
of the Skeletal System
7.1.3.1
Skull
7.1.3.1.1
Sutures
7.1.3.1.2
Fontanels
7.1.3.1.3
Cranial Bones
7.1.3.1.3.1 Frontal Bone
7.1.3.1.3.2 Parietal Bones
7.1.3.1.3.3 Temporal Bones
7.1.3.1.3.4 Occipital Bone
7.1.3.1.3.5 Sphenoid Bone
7.1.3.1.3.6 Ethmoid Bone
7.1.3.1.3.7 Facial Bones
7.1.3.1.3.8 Nasal Bones
7.1.3.1.3.9 Maxillae
7.1.3.1.3.10 Paranasal Sinuses
7.1.3.1.3.11 Zygomatic Bones
7.1.3.1.3.12 Mandible
7.1.3.1.3.13 Lacrimal Bones
7.1.3.1.3.14 Palatine Bones
7.1.3.1.3.15 Inferior Nasal Conchae
7.1.3.1.3.16 Vomer
7.1.3.1.4
Orbits
7.1.3.1.5
Foramina
7.1.3.2
Hyoid
Bone
7.1.3.3
Vertebral
Column
7.1.3.3.1
Divisions
7.1.3.3.2
Normal Curves
7.1.3.3.3
Typical
Vertebra
7.1.3.3.4
Cervical
Region
7.1.3.3.5
Thoracic
Region
7.1.3.3.6
Lumbar Region
7.1.3.3.6.1
Lumbar vertebrae Illustration # 8 Illustration # 9 Netter 27
7.1.3.3.6.1.1
The
lumbar vertebrae, five in number, are large and massive because of their
weight-bearing function and are chiefly characterized by having no foramen in
their transverse processes and no articular facets on their vertebral bodies.
The lumbar curve is convex anteriorly with interposing fibrocartilaginous discs
for mobility.
7.1.3.3.6.1.2
The
spinal foramen in the lumbar region is triangular though smaller than in the
cervical region. The body of each lumbar vertebra is narrower from before
backward and wider from side to side. It is slightly taller anteriorly than
posteriorly, and shows more concavity above than below. The pedicles arise from
either side of the upper portion of the vertebral body. They are short, strong
and proceed directly backward with grooves above and below forming the superior
and inferior vertebral notches. These join to form foraminae for the exit of
the spinal nerves.
7.1.3.3.6.1.3 Springing from the pedicles, are the
short, broad and powerful laminae, which meet in the midline to form the
spinous process. This process is thick and. broad and directed backward in an
almost horizontal direction.
7.1.3.3.6.1.4
At the
base of the laminae are the articular facets. The superior articular facet is
concave and directed medially and backward whereas the inferior articular facet
is directed forward and laterally. The transverse processes, which arise from
the pedicles, are comparatively slender and are situated in front of the
articular 27 processes.
7.1.3.3.6.1.5
On the
posterior superior aspect of the articular facets are slight elevations called
mammillary processes. A similar elevation on the posterior aspect of the
transverse process is called the accessory process.
7.1.3.3.6.1.6
The
intervertebral disc, a fibrocartilaginous structure, acts as a cushion between
the adjacent vertebral bodies. It varies in size and shape and conforms to the
corresponding vertebral body. These discs are thickest in the lumbar region,
increasing in size as they approach the sacrum.
7.1.3.3.6.1.7 The fibrocartilaginous tissue is concentrically arranged with a thick and tough outer layer called the annulus fibrosus and a soft gelatinous pulpy substance called the nucleus pulposus, which are the remains of the notochord. The intervertebral discs are particularly poor in blood supply.
7.1.3.3.7
Sacrum and
Coccyx
7.1.3.3.7.1 Sacrum Illustration # 7 Netter 29
7.1.3.3.7.1.1
In the
adult, the sacrum articulates with the fifth lumbar vertebra, the coccyx and
the two hipbones. It is large, triangular and wedge-shaped and forms the
posterior wall of the pelvis. Its base is oval-shaped and articulates with the
inferior aspect of the body of the fifth lumbar vertebra, forming the prominent
sacral vertebral angle. On either side of the sacrum are triangular areas
called the alae. Just below and to either side of the base is a ridge forming
the sacral portion of the pelvic brim. The lower portion of the sacrum, the
apex, articulates with the coccyx.
7.1.3.3.7.1.2
The
pelvic surface of the sacrum is concave in both dimensions, vertically and
horizontally. At its middle, it is grooved with four transverse ridges, which
mark the points of fusion of the original five vertebrae. There are four
anterior sacral foraminae on either side of the ridges, which permit passage of
the anterior division of the sacral nerve and the lateral sacral arteries.
7.1.3.3.7.1.3 A cross-sagittal section of the sacrum demonstrates the sacral canal which en- closes the lower portion of the dura and contents, the roots of the cauda equina, the filum terminale externum, fat, areolar tissue, blood vessels and fine nerve filaments.
7.1.3.3.8
Thorax
7.1.3.3.8.1 Sternum
7.1.3.3.8.2 Ribs
8 Skeletal System-Appendicular Skeleton*
8.1
Pectoral
(Shoulder) Girdle
8.2
Upper
Extremity
8.3
Pelvic
(Hip) Girdle
8.4
Lower
Extremity
8.4.1
8.4.2
Femur
8.4.3
Patella
8.4.4
Tibia
and Fibula
8.4.4.1
Tibia
8.4.4.1.1 The tibia, or shinbone, is the larger, medial bone of the leg (Figure 8-12). It bears the major portion of the weight of the leg. The tibia articulates at its proximal end with the femur and fibula, and at its distal end with the fibula of the leg and talus bone of the ankle.
8.4.4.1.2 Figure Photograph of right tibia and fibula (a) Anterior View (b) Posterior View
8.4.4.1.2.1 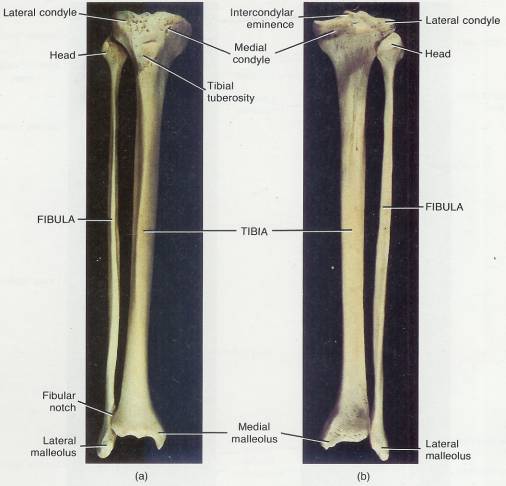
8.4.4.1.3 The proximal end of the tibia is expanded into a lateral condyle and a medial condyle. These articulate with the condyles of the femur. The inferior surface of the lateral condyle articulates with the head of the fibula. The slightly concave condyles are separated by an upward projection called the intercondylar eminence. The tibial tuberosity on the anterior surface is a point of attachment for the patellar ligament.
8.4.4.1.4 The medial surface of the distal end of the tibia forms the medial malleolus (mal-LĒ-ō-lus). This structure articulates with the talus bone of the ankle and forms the prominence that can be felt on the medial surface of your ankle. The fibular notch articulates with the fibula.
8.4.4.2
Fibula
8.4.4.2.1
The fibula is parallel and lateral
to the tibia. It is considerably smaller than the tibia and is non-weight
bearing. The head of
the fibula, the proximal end, articulates with the inferior surface of the
lateral condyle of the tibia below the level of the knee joint. The distal end
has a projection called the lateral
malleolus that articulates with the talus bone of the ankle. This forms
the prominence on the lateral surface of the ankle. The inferior portion of the
fibula also articulates with the tibia at the fibular notch. A fracture of the
lower end of the fibula with injury to the tibial articulation is called a Pott's fracture.
8.4.5
Tarsals,
Metatarsals, and Phalanges
8.4.5.1
Tarsus
(Tarsals) (7) (tahr’sus)
8.4.5.1.1
The tarsus is
a collective designation for the seven bones of the ankle called tarsals
(Figure 8-13). The term tarsos pertains to a broad, flat surface.
8.4.5.1.2
Figure 8-13
Photograph of the right foot in superior view
8.4.5.1.2.1
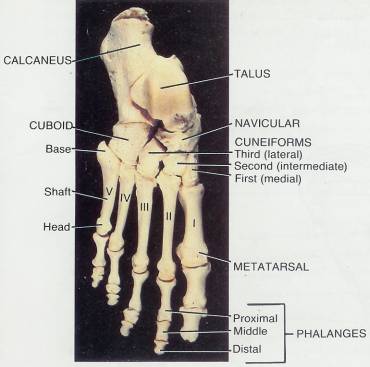
8.4.5.1.3 The talus and calcaneus (kal-KĀ-nē-us) are located on the posterior part of the foot. The anterior part contains the cuboid, navicular, and three cuneiform (cuneiform = wedge-shaped) bones called the first (medial), second (intermediate), and third (lateral) cuneiform. The talus, the uppermost tarsal bone, is the only bone of the foot that articulates with the fibula and tibia. It is surrounded on one side by the medial malleolus of the tibia and on the other side by the lateral malleolus of the fibula. During walking, the talus initially bears the entire weight of the body. About half the weight is then transmitted to the calcaneus. The remainder is transmitted to the other tarsal bones. The calcaneus, or heel bone, is the largest and strongest tarsal bone.
8.4.5.2 Metatarsals
8.4.5.2.1 The metatarsus consists of five metatarsal bones numbered I to V from the medial to lateral position. Like the metacarpals of the palm of the hand, each metatarsal consists of a proximal base, a shaft, with a distal head. The metatarsals articulate proximally with the first, second, and third cuneiform bones and with the cuboid. Distally, they articulate with the proximal row of phalanges. The first metatarsal is thicker than the others because it bears more weight.
8.4.5.3 Phalanges
8.4.5.3.1
The phalanges
of the foot resemble those of the hand both in number and arrangement.
Each also consists of a proximal base,
a middle shaft,
and a distal head.
The hallux (great or big
toe), has two large, heavy phalanges called proximal and distal phalanges. The
other four toes each have three phalanges-proximal, middle, and distal.
8.4.6
Arches
of the Foot
8.4.6.1
The
bones of the foot are arranged in two arches (Figure 8-14). These arches enable
the foot to support the weight of the body, provide an ideal distribution of body
weight over the hard and soft tissues of the foot, and provide leverage while
walking.
8.4.6.2
Figure
8-14 Arches of the right foot in lateral View
8.4.6.2.1
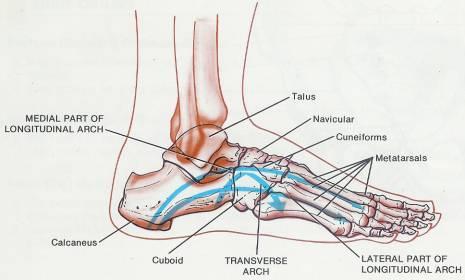
8.4.6.3
The
arches are not rigid. They yield as
weight is applied and spring back when the weight is lifted, thus helping to
absorb shocks.
8.4.6.4
The
longitudinal arch has two parts. Both consist of tarsal and metatarsal bones
arranged to form an arch from the anterior to the posterior part of the foot.
The medial (inner) part of the longitudinal arch originates at the calcaneus.
It rises to the talus and descends through the navicular, the three cuneiforms,
and the heads of the three medial metatarsals. The talus is the keystone of
this arch. The lateral (outer) part of the longitudinal arch also begins at the
calcaneus. It rises at the cuboid and descends to the heads of the two lateral
metatarsals. The cuboid is the keystone of this arch.
8.4.6.5
The
transverse arch is formed by the navicular, three cuneiforms, cuboid, and the
bases of the five metatarsals.
8.4.7
Clinical
Application: Flatfoot, Clawfoot, and Bunions
8.4.7.1
The
bones composing the arches are held in position by ligaments and tendons. If
these ligaments and tendons are weakened, the height of the medial longitudinal
arch may decrease or “fall”. The result is flat foot.
8.4.7.2
Clawfoot
is a condition in which the medial longitudinal arch is abnormally elevated. It
is frequently caused by muscle imbalance, such as may result from
poliomyelitis.
8.4.7.3
A
bunion (hallux valgus; valgus bent outward) is a deformity of the great toe.
Although the condition may be inherited, it is typically caused by wearing
tightly fitting shoes and is characterized by lateral deviation of the great
toe and medial displacement of metatarsal 1. Arthritis of the first
metatarsophalangeal joint may also be a predisposing factor. The condition
produces inflammation of the bursae (fluid-filled sacs at the joints), bone
spurs, and calluses.
8.5
Female
and Male Skeletons
9 Articulations
9.1 Introduction
9.1.1 Bones are too rigid to bend without causing damage. Fortunately, the skeletal system consists of many separate bones, most of which are held together at joints by flexible connective tissue. All movements that change the positions of the bone parts of the body occur at joints. You can understand the importance of joints if you imagine how a cast over the knee joint prevents flexing the leg or how a splint on a finger limits the ability to manipulate small objects.
9.1.2 An articulation (joint) is a point of contact between bones between cartilage and bones, or between teeth and bones. The scientific study of joints is referred to as Arthrology (ar-THROL-ō-jē; arthro=joint; logos=sturdy of). The joint’s structure determines how it functions. Some joints permit no movement, other permit slight movement, and still others afford considerable movement. In general, the closer the fit at the point of contact, the stronger the joint. At tightly fitted joints, however, movement is restricted. The looser the fit, the greater the movement. Unfortunately, loosely fitted joints are prone to dislocation. Movement at joints is also determined by the structure (shape) of the articulation bones, the flexibility (tension or tautness) of the connective tissue ligaments and joint capsules that bind the bones together, and the position of ligaments, muscles, and tendons.
9.2 Classification of Joints
9.2.1 Functional
9.2.1.1 Introduction
9.2.1.1.1 The functional classification of joints takes into account the degree of movement they permit.
9.2.1.2 Synarthroses (sin-ar-THRŌ-sēz))
9.2.1.2.1 Immovable
9.2.1.3 Amphiarthroses (am-fē –ar-THRŌ-sēz)
9.2.1.3.1 Slightly moveable
9.2.1.4 Diarthroses (dī-ar-THRŌ-sēz)
9.2.1.4.1 Freely movable
9.2.2 Structural
9.2.2.1 The structural classification of joints is based on the presence or absence of a synovial (joint) cavity (a space between the articulating bones) and the kind of connective tissue that binds the bones together. Structurally, joints are classified as Fibrous Joints, in which there is no synovial cavity and the bones are held together by fibrous connective tissue. Cartilaginous Joints have no synovial cavity and the bones are held together by cartilage. Synovial Joints have a synovial cavity and the bones forming the joint are united by a surrounding articular capsule and frequently by accessory ligaments (described later). We will discuss the joints of the body based upon their structural classification, but with reference to their functional classification as well.
9.3 Fibrous Joints
9.3.1 Fibrous joints lack a synovial cavity, and the articulating bones are held very closely together by fibrous connective tissue. They permit little or no movement. The three types of fibrous joints are Sutures, Syndesmoses, and gomphoses.
9.3.2 Suture
9.3.2.1
9.3.3 Syndesmosis (Sin’-dez-MŌ-sis)
9.3.3.1 This is a fibrous joint in which the uniting fibrous connective tissue is present in a much greater amount than in a suture, but the fit between the bones is not quite as tight.
9.3.3.2 The fibrous connective forms an interosseous membrane or ligament.
9.3.3.3 A Syndesmosis is slightly movable because the bones are separated more than in a suture and some flexibility is permitted by the interosseous membrane or ligament.
9.3.3.4 Syndesmoses are functionally classified as amphiarthrotic and typically permit slight movement.
9.3.3.5 An example of a Syndesmosis is the distal articulation of the fibia and fibula
9.3.4 Gomphosis
9.3.4.1
9.4 Cartilaginous Joints
9.4.1
9.4.2 Synchondrosis
9.4.2.1
9.4.3 Symphysis
9.4.3.1
9.5 Synovial Joints (si-NŌ-vē-al)
9.5.1 Structure
9.5.1.1
A
joint in which there is a space between articulating bones is called a synovial
joint. The space is called a synovial (joint) cavity. Because of this cavity
and because of the arrangement of the articular capsule and accessory
ligaments, synovial joints are freely movable. Thus, synovial joints are
functionally classified as diarthrotic.
9.5.1.2
Synovial
joints are different from the other joint types in the following ways.
9.5.1.2.1 Synovial Joint Cavity
9.5.1.2.1.1 When a joint has a space between its cartilaginous end surfaces, it is called a Synovial (si-NŌ-vē-al) joint.
9.5.1.2.1.2 This space is called a Synovial (joint) cavity. This cavity along with the articular capsule and its accessory ligaments allows Synovial joints to be freely movable.
9.5.1.2.2 Hyaline (Articular) Cartilage
9.5.1.2.2.1 The hyaline (articular) cartilage covering the end surfaces of bones is another distinctive feature of Synovial joints. It provides a smooth, relatively friction free surface around which bones can move unimpeded.
9.5.1.2.3 Articular Capsule
Back Table of Contents References
9.5.1.2.3.1 The articulating bones are united by this sleeve like capsule, which is composed of two layers as follows.
9.5.1.2.3.1.1 Fibrous Capsule
9.5.1.2.3.1.1.1 The fibrous capsule is the outermost layer of the articular capsule.
9.5.1.2.3.1.1.2 The fibrous capsule is composed of dense connective (collagenous) tissue.
9.5.1.2.3.1.1.3 It is attached to the periosteum of articulating bones at variable distance from the edge of the articular cartilage.
9.5.1.2.3.1.1.4 It is flexible so that it permits movement at the joint. It has great tensile strength so that it resists dislocation.
9.5.1.2.3.1.1.5 Some fibrous capsules have outer layers of its fibers arranged in parallel bundles called ligaments, which are individually named.
9.5.1.2.3.1.1.6 Ligaments resist repetitive strain and bind bone to bone.
9.5.1.2.3.1.2 Synovial Membrane
9.5.1.2.3.1.2.1 This is the innermost layer of the articular capsule and is another distinguishing feature of Synovial joints.
9.5.1.2.3.1.2.2 It is composed of a variable amount of adipose (fat) and loose connective tissue with elastic fibers.
9.5.1.2.3.1.2.3 The Synovial membrane secretes Synovial fluid (SF), which provides both nourishment to the articular cartilage and lubrication to the joint.
9.5.1.2.3.1.2.4 Phagocytic cells found in the Synovial fluid remove microbes and dead cells (debris) caused by wear and tear on the joint.
9.5.1.2.3.1.2.5 The Synovial fluid consists of hyaluronic acid and an interstitial fluid produced from blood plasma.
9.5.1.2.3.1.2.6 Synovial fluid is similar in consistency to uncooked egg white, which is viscous when there is no joint movement and less viscous with increased joint movement.
9.5.1.2.3.1.2.7 There is just enough Synovial fluid in the joint to form a thin viscous layer over the joint surfaces. The thin film reduces friction and supplies nutrients and removes metabolic waste from the cartilage cells of the articulate cartilage. The amount of synovial fluid will vary from joint to joint. The amount of Synovial fluid in a large joint such as the knee is about 3.5 ml (0.1 oz).
9.5.1.3 Crackling Sound
Back Table of Contents References
9.5.1.3.1 When some synovial joints are pulled apart, they make a crackling sound. As space is created in a joint, negative pressure develops in the synovial fluid driving out carbon dioxide. As a result a gas bubble develops. As the opposing articular surfaces are pulled apart further and then abruptly, separate the pressure within the joint exceeds that in the bubble and the bubble collapses, producing a crackling noise. The collapse of the large bubble creates a series of smaller bubbles, which gradually go back into solution. Until the small bubbles disappear and the gas is completely dissolved, the joint cannot be cracked again. Increased congruence of joint surfaces such as those found between the phalanges and metacarpals, crack more easily than the joint surfaces that are less congruent.
9.5.1.4 Accessory ligaments
9.5.1.4.1 There are two kinds of accessory ligaments (ligare= to bind) one which is on the outside of the joint capsule is named the extracapsular ligament and another is named the intracapsular ligament because it is on the inside of a joint capsule.
9.5.1.4.2 Extracapsular Ligaments
9.5.1.4.2.1 These accessory ligaments are outside the articular capsule. An example would be the fibular collateral ligament of knee joint.
9.5.1.4.3 Intracapsular Ligaments
9.5.1.4.3.1 These accessory ligaments are within the articular capsule but excluded from the synovial cavity by reflections of a synovial membrane. The cruciate ligaments within the knee joint are an example.
9.5.1.5 Articular Discs (Menisci)
9.5.1.5.1 The menisci are pads of fibrocartilage between the articular surfaces of the bone. They attach to the margins of the fibrous capsule. The menisci subdivide the space between the joint into two separate areas and allow two bones of different shapes to fit snugly. This helps maintain the stability of the joint while also directing the synovial fluid to areas of greatest friction.
9.5.1.6 Bursae
Back Table of Contents References
9.5.1.6.1 Bursae help reduce friction between moving parts. Bursae are saclike structures whose walls consist of connective tissue similar to the fibrous tissue around the joint capsule. Bursae are lined with a synovial membrane and filled with fluid similar to synovial fluid. Bursae cushion movement of one part of a body over another and repetitive stress can inflame them. This inflammation is called bursitis. Bursae are found between the following structures.
9.5.1.6.1.1 Skin and bone in places where the skin rubs over the bone.
9.5.1.6.1.2 Tendons and bones.
9.5.1.6.1.3 Muscles and bones
9.5.1.6.1.4 Ligaments and bones
9.5.1.7 Joint Stability
9.5.1.7.1 Joint stability requires that bones remain in contact with each other. Bone to bone contact is facilitated by several factors as follows;
9.5.1.7.2 Fit of articulating bones
9.5.1.7.2.1 In the hip joint, for example, the head of the femur and the acetabulum of the coxal (hip) bone interlock with one another.
9.5.1.7.3 Strength and tension (tautness) of the joint ligaments
9.5.1.7.3.1 The thick ligaments of the coxal (hip) joint help provide stability to the articulating bones.
9.5.1.7.4 Tension and arrangement of muscles around the joint
9.5.1.7.4.1 The knee joint, for example, is surrounded by a fibrous capsule which is formed primarily from the tendinous expansions of the muscles acting on the joint.
9.5.1.8 Medical Tests, Procedures, medical conditions and definitions
Back Table of Contents References
9.5.1.8.1 Arthrocentesis (ar-thrō-sen-TĒ-sis; arthro= joint; centesis= to puncture) (joint tap)
9.5.1.8.1.1 Diagnostic and or Therapeutic value
9.5.1.8.1.1.1 This procedure removes a sample of synovial fluid for analysis. The sample once analyzed helps diagnose the cause of joint swelling, inflammation, and or pain.
9.5.1.8.1.1.2 Fluid is drained from the joint capsule. To decrease the pressure within the joint capsule caused by a build up of synovial fluid, blood, and/or pus.
9.5.1.8.1.1.3 Local anesthesia or steroid medication is injected into the joint capsule. This procedure helps reduce the pain of arthritis.
9.5.1.8.1.1.4 Abnormal joint fluid is drained from the capsule and the joint is cleansed by saline irrigations.
9.5.1.8.1.2 Procedure
9.5.1.8.1.2.1 Antiseptic solution is applied to the skin over the joint for cleansing.
9.5.1.8.1.2.2 An aesthetic is injected into the area.
9.5.1.8.1.2.3 A sample of synovial fluid is obtained by inserting a needle into the synovial joint cavity.
9.5.1.8.1.2.4 Fluid analysis is completed for cell content, which includes urate and other crystals.
9.5.1.8.1.2.5 Color and viscosity are also noted.
9.5.1.8.1.2.6 A sample is submitted for microbial culture and chemical analysis, which includes glucose, protein, and enzymes.
9.5.1.8.2 Artificial Ligaments and Tendons
Back Table of Contents References
9.5.1.8.2.1 These ligaments and tendons are used to replace ligaments and tendons, which are damaged. An example of this would be the carbon fiber implant. Carbon fibers are coated with a plastic called polylactic acid. Ligaments and tendons are then reinforced by these coated fibers, which are sewn in and around them. The polylactic acid is absorbed within about two weeks by the body and the carbon fibers eventually fracture. As the artificial fibers either are absorbed by the body or break apart, they are surrounded by collagen produced by fibroblasts.
9.5.1.8.3 Torn cartilage
9.5.1.8.3.1 Articular disks (menisci) are torn (torn cartilage) usually during athletic activity. Surgical removal (meniscectomy) is necessary to avoid wear and tear, which may result in arthritis. This surgery, which once involved removal of much if not all of the cartilage, also damaged healthy tissue and required extensive time-consuming rehabilitation. Arthroscopy reduces the damage to tissue and shortens rehabilitation time.
9.5.1.8.4 Arthroscopy (ar-THROS-kō-pē; arthro= joint; skopein= to view)
9.5.1.8.4.1 An arthroscope is a lighted instrument about the diameter of the pencil, which is used to examine the interior of the joint usually a knee.
9.5.1.8.4.2 Diagnostic value
9.5.1.8.4.2.1 Arthroscopy provides internal Assessment of the damage following a knee injury.
9.5.1.8.4.2.2 Arthroscopy helps remove torn cartilage, and repair cruciate ligaments within the joint capsule.
9.5.1.8.4.2.3 Tissue samples can be obtained and analyzed.
9.5.1.8.4.2.4 Disease progression and set the effects can be monitored.
9.5.1.8.4.2.5 Surgical procedures can be planned.
9.5.1.8.4.3 Procedure
9.5.1.8.4.3.1 A local or general anesthetic is administered.
9.5.1.8.4.3.2 A quarter inch incision is made and the arthroscope is inserted.
9.5.1.8.4.3.3 A tube for the saline solution is inserted in another small incision. The arthroscope itself can also introduce saline solution.
9.5.1.8.4.3.4 An additional small incision allows for the insertion of an instrument that shaves off, reshapes the damaged cartilage, and removes the shaved cartilage along with the salt solution.
9.5.1.8.4.3.5 A video camera can be attached to the arthroscope for real time screen projection or later viewing.
9.5.1.8.4.3.6 Because of the small incision is recovery time is much improved over methods that are more conventional.
9.5.1.8.4.3.7 The patient may wear a cast or split for several days depending on the extent of the procedure
9.5.2 Movements
9.5.2.1 The movements permitted at synovial joints are limited by several factors.
9.5.2.2 Structure (Shape) of the Articulating Bones
9.5.2.2.1 The shape of the articulating bones and how they fit together will determine the movements permitted at that synovial joint.
9.5.2.3 Tension of Ligaments
9.5.2.3.1 The different components of a fibrous capsule are tense only when the joint is in certain positions. Tense ligaments not only restrict the range of movement but also direst the movement of the articulating bones with respect to each other. In the knee joint, for example, the major ligaments are lax when the knee is bent but tense when the knee is straightened. Also, when the knee is straightened, the surfaces of the articulating bones are in fullest contact with each other.
9.5.2.4 Muscle Arrangement and Tension
9.5.2.4.1 How the muscles are arranged and the tension they place on the join, which reinforce the restraint placed on a joint by ligaments is another factor which restricts movement at a synovial joint.
9.5.2.5 Apposition of Soft Parts
9.5.2.5.1 This may limit mobility. For example, during bending at the elbow, the anterior surface of the forearm is pressed against the anterior surface of the arm.
9.5.2.6 Gliding
9.5.2.6.1 A gliding movement is the simplest kind that can occur at a joint. One surface moves back and forth and from side to side over another surface without angular or rotary motion. Some joints that glide are those between the carpals and between the tarsals. The heads and tubercles of the ribs glide on the bodies and transverse processes of the vertebrae.
9.5.2.7 Angular
9.5.2.7.1 Increase or decrease the angle between bones. Among the angular movements are flexion, extension, abduction, and adduction.
9.5.2.7.2 Flexion
9.5.2.7.2.1 Decreases the angle between the surfaces of the articulating bones. Examples of flexion include bending the head forward (the joint is between the occipital bone and the atlas), bending the elbow, and bending the knee.
9.5.2.7.3 Extension
9.5.2.7.3.1 Involves an increase in the angle between the surfaces of the articulating bones. Extension restores a body part to its anatomical position after it has been flexed. Examples of extension are returning the head to the anatomical position after flexion, straightening the arm after flexion, and straightening the leg after flexion. Continuation of extension beyond the anatomical position, as in bending the head backward, is called hyperextension.
9.5.2.7.4 Abduction
9.5.2.7.4.1 Movement of a bone away from the midline of the body. An example of abduction is moving the arm upward and away from the body until it is held straight out at right angles to the chest. With the fingers and toes, however, the midline of the body is not used as the line of reference. Abduction of abduction of the fingers (not the thumb) is a movement away from an imaginary oline drawn through the middle finger; in other words, is is spreading the fingers. Abduction of the thumb moves the thumb away from the plane of the palm at a right angle to the palm. Abduction of the toes is relative to an imaginary line drawn through the second toe.
9.5.2.7.4.2 Further Reading
9.5.2.7.4.2.1 Adduction and Abduction
9.5.2.7.5 Adduction
9.5.2.7.5.1 Movement of a part toward the midline of the body. An example of adduction is returning the arm to the side after abduction. As in abduction, adduction of the fingers (not the thumb) is relative to the middle finger, and adduction of the toes is relative to the second toe. In adduction of the thumb, the thumb moves toward the plane of the palm at a right angle to the palm.
9.5.2.7.5.2 Further Reading
9.5.2.7.5.2.1 Adduction and Abduction
9.5.2.8 Rotation
9.5.2.8.1 Movement of a bone around its own longitudinal axis. During rotation, no other motion is permitted.
9.5.2.8.1.1 Medial Rotation
9.5.2.8.1.1.1 Anterior surface of a bone or extremity moves toward the midline.
9.5.2.8.1.2 Lateral Rotation
9.5.2.8.1.2.1 Anterior surface moves away from the midline.
9.5.2.8.2 We rotate the atlas around the dens of the axis when we shake the head from side to side. Rotation of the humerus turns the anterior surface of the humerus to face wither medially or laterally.
9.5.2.9 Circumduction
9.5.2.9.1 Movement in which the distal end of a bone moves in a circle while the proximal end remains stable. The bone describes a cone in the air. Circumduction typically involves flexion, abduction, adduction, extension, and rotation. It involves a 360° rotation. An example is moving the outstretched arm in a circle toward up to pitch a ball.
9.5.2.10 Special
9.5.2.10.1 Special movements are those found only at the joints indicated
9.5.2.10.1.1 Inversion
9.5.2.10.1.1.1 Movement of the sole of the foot inward (medially) so that the soles face toward each other.
9.5.2.10.1.1.2 Further Reading
9.5.2.10.1.1.2.1 Inversion
9.5.2.10.1.2 Eversion
9.5.2.10.1.2.1 Movement of the sole outward (laterally) so that the soles face away from each other
9.5.2.10.1.2.2 Further Reading
9.5.2.10.1.2.2.1 Eversion
9.5.2.10.1.3 Dorsiflexion
9.5.2.10.1.3.1 Involves bending of the foot in the direction of the dorsum (upper surface) of the foot.
9.5.2.10.1.4 Plantar Flexion
9.5.2.10.1.4.1 Involves bending the foot in the direction of the plantar surface (sole) of the foot.
9.5.2.10.1.4.2 Further Reading
9.5.2.10.1.4.2.1 Flexion and Extension
9.5.2.10.1.5 Protraction
9.5.2.10.1.5.1 Movement of the mandible or shoulder girdle forward on a plane parallel to the ground. Thrusting the jaw outward is protraction of the mandible. Bring your arms forward until the elbows touch requires protraction of the clavicle or shoulder girdle.
9.5.2.10.1.6 Retraction
9.5.2.10.1.6.1 Movement of a protracted part of the body backward on a plane parallel to the ground. Pulling the lower jaw back in line with the upper jaw is retraction of the mandible.
9.5.2.10.1.7 Supination
9.5.2.10.1.7.1 Movement of the forearm in which the palm of the hand is turned anterior or superior. To demonstrate supination, flex your forearm at the elbow to prevent rotation of the humerus in the shoulder joint.
9.5.2.10.1.7.2 Further Reading
9.5.2.10.1.7.2.1 Supination and Pronation
9.5.2.10.1.7.2.2
9.5.2.10.1.8 Pronation
9.5.2.10.1.8.1 Movement of the forearm in which the palm is turned posterior or ingerior.
9.5.2.10.1.8.2 Further Reading
9.5.2.10.1.8.2.1 Supination and Pronation
9.5.2.10.1.9 Elevation
9.5.2.10.1.9.1 Upward movement of a part of the body. You elevate your mandible when you close your mouth.
9.5.2.10.1.10 Depression
9.5.2.10.1.10.1 Downward movement of a part of the body. You depress your mandible when you open your mouth. The shoulders can also be elevated and depressed.
9.5.3 Types of Synovial Joints
Back Table of Contents References
9.5.3.1 Types
9.5.3.1.1 The shape of the articulating bones determines the type of movement that can be performed. The structure of synovial joints although similar differ in articular shape and can be divided into six sub-types as follows; Gliding, Hinge, Pivot, Ellipsoidal, Saddle, and Ball and Socket.
9.5.3.2 Gliding
9.5.3.2.1 The bones of gliding joints (arthrodia (ar-THRŌ-dē-a)) form a flat surface so that only side-to-side and back and fourth movements are possible. The ligaments or adjacent bones limit twisting and rotation movements. Gliding joints are nonaxial because there's no axis around which they move.
9.5.3.2.2 Movements
9.5.3.2.2.1 Gliding
9.5.3.2.3 Examples
9.5.3.2.3.1 Joints between the carpal bones
9.5.3.2.3.2 Tarsal bones
9.5.3.2.3.3 Sternum and clavicle
9.5.3.2.3.4 Scapula and clavicle
9.5.3.3 Hinge
9.5.3.3.1 The hinge or ginglymus (JIN-gli-mus) joint is one in which the concave surface of one bone fits into the convex surface of another bone. This joint is monaxial or uniaxial because its movement is primarily a single plane. A door hinge is a good example of this type of joint.
9.5.3.3.2 Movements
9.5.3.3.2.1 Flexion
9.5.3.3.2.2 Extension
9.5.3.3.3 Examples
9.5.3.3.3.1 Elbow
9.5.3.3.3.2 Ankle
9.5.3.3.3.3 Interphalangeal joints
9.5.3.4 Pivot
Back Table of Contents References
9.5.3.4.1 A pivot or trochoid (TRŌ-koyd) is formed between the rounded, pointed, or conical surface of one bone, which articulates within a ring, formed partly by another bone and partly by ligament. This joint is monaxial in that the primary movement permitted is rotation.
9.5.3.4.2 Movements
9.5.3.4.2.1 Supination and pronation of the palms
9.5.3.4.2.2 Rotation of the head from side to side
9.5.3.4.3 Examples
9.5.3.4.3.1 Atlas and axis (atlantoaxial)
9.5.3.4.3.2 Proximal ends of the Radius and ulna
9.5.3.5 Ellipsoidal
9.5.3.5.1 An ellipsoidal or condyloid (KON-di-loyd) joint, an oval-shaped condyle of one bone fits into an elliptical cavity of another bone. Since the joint permits side-to-side and back-and-forth movements, it is biaxial. The joint at the wrist between the radius and carpals is ellipsoidal. The movement permitted by such a joint is illustrated when you flex and extend and abduct and adduct and circumduct the wrist.
9.5.3.5.2 Movements
9.5.3.5.2.1 Flex and extend and abduct and adduct and circumduct the wrist.
9.5.3.5.3 Examples
9.5.3.5.3.1 Wrist
9.5.3.5.3.2 Metatarsophalangeal Joints (MP)
9.5.3.6 Saddle (Reciprocal Reception)
9.5.3.6.1 In a saddle or sellaris (sel-A-ris) joint, the articular surface of one bone is saddle-shapped and the articular surface of the other bone is shaped like a rider sitting in the saddle. Essentially, the saddle joint is a modified ellipsoidal joint in which the movement is somewhat freer. Movements at a saddle joint are die to side and back and forth. Thus, the joint is biaxial.
9.5.3.6.2 Movements
9.5.3.6.2.1 Side to side and back and forth
9.5.3.6.3 Examples
9.5.3.6.3.1 Trapezium of the carpus and the metacarpal of the thumb
9.5.3.6.3.2 Calcaneocuboid Joint of foot
9.5.3.7 Ball and Socket
9.5.3.7.1 A ball and socket or spheroid (SFĒ-royd) joint consists of a ball-like surface of one bone fitted into a cuplike depression of another bone. Such a joint permits triaxial movement, or movement in three planes of motion: flexion-extension, abduction-adduction, and rotation-Circumduction. The range of movement at a ball-and-socket joint is illustrated by Circumduction of the arm.
9.5.3.7.2 Movements
9.5.3.7.2.1 Flexion-extension,
9.5.3.7.2.2 Abduction-adduction
9.5.3.7.2.3 Rotation-Circumduction
9.5.3.7.3 Examples
9.5.3.7.3.1 Shoulder Joint
9.5.3.7.3.2 Coxal (hip) Joint
9.5.3.7.3.3 Talocalcaneonavicular (TCN) of the foot
9.5.4 Discussion of Selected Synovial Joints
Back Table of Contents References
9.5.4.1 Humeroscapular (Shoulder) Joint
9.5.4.1.1
9.5.4.1.2 Articular Capsule
9.5.4.1.3 Coracohumeral Ligaments
9.5.4.1.4 Transverse Humeral Ligament
9.5.4.1.5 Transverse Humeral Ligament
9.5.4.1.6 Glenoid Labrum
9.5.4.1.7 Bursae
9.5.4.1.7.1 Subscapular Bursa
9.5.4.1.7.1.1
9.5.4.1.7.2 Subdeltoid Bursa
9.5.4.1.7.2.1
9.5.4.1.7.3 Subacromial Bursa
9.5.4.1.7.3.1
9.5.4.1.7.4 Subcoracoid Bursa
9.5.4.1.7.4.1
9.5.4.2 Coxal (Hip) Joint
Back Table of Contents References
9.5.4.2.1
9.5.4.2.2 Articular Capsule
9.5.4.2.2.1
9.5.4.2.3 Iliofemoral Ligament
9.5.4.2.3.1
9.5.4.2.4 Pubofemoral Ligament
9.5.4.2.4.1
9.5.4.2.5 Ischiofemoral Ligament
9.5.4.2.5.1
9.5.4.2.6 Capitate Ligament (Ligament of the head of the femur)
9.5.4.2.6.1
9.5.4.2.7 Acetabular Labrum
9.5.4.2.7.1
9.5.4.2.8 Transverse Ligament of the Acetabulum
9.5.4.2.8.1
9.5.4.2.9 Dislocation of the Hip
9.5.4.2.9.1
9.5.4.3 Tibiofemoral (Knee) Joint
Back Table of Contents References
9.5.4.3.1
9.5.4.3.2 Articular Capsule
9.5.4.3.2.1
9.5.4.3.3 Medial and Lateral Patellar Retinacula
9.5.4.3.3.1
9.5.4.3.4 Patellar Ligament
9.5.4.3.4.1
9.5.4.3.5 Oblique Popliteal Ligament
9.5.4.3.5.1
9.5.4.3.6 Arcuate Popliteal Ligament
9.5.4.3.6.1
9.5.4.3.7 Tibial Collateral Ligament
9.5.4.3.7.1
9.5.4.3.8 Fibular Collateral Ligament
9.5.4.3.8.1
9.5.4.3.9 Intraarticular Ligaments
9.5.4.3.9.1 Anterior Cruciate Ligament
9.5.4.3.9.1.1
9.5.4.3.9.2 Posterior Cruciate Ligament
9.5.4.3.9.2.1
9.5.4.3.10 Articular Discs
9.5.4.3.10.1 Medial Meniscus
9.5.4.3.10.1.1
9.5.4.3.10.2 Lateral Meniscus
9.5.4.3.10.2.1
9.5.4.3.11 Bursae
Back Table of Contents References
9.5.4.3.11.1 Anterior Bursae
9.5.4.3.11.1.1
9.5.4.3.11.2 Medial Bursae
9.5.4.3.11.2.1
9.5.4.3.11.3 Lateral Bursae
9.5.4.3.11.3.1
9.5.4.3.12 Knee Injuries
9.5.4.3.12.1 Rupture of Tibial Collateral Ligament, Tearing of Anterior Cruciate & Medial Meniscus
9.5.4.3.12.1.1
9.5.4.3.12.2 Swollen Knee
9.5.4.3.12.2.1
9.5.4.3.12.3 Dislocated Knee
9.5.4.3.12.3.1
9.5.5 Exhibit 9-3
9.5.5.1

9.5.5.2

9.6 Disorders: Homeostatic Imbalance
Back Table of Contents References
9.6.1 Rheumatism (rheumat=Subject to flux)
9.6.1.1 Refers to any painful state of the supporting structures of the body- its bones, ligaments, joints, tendons, or muscles. Arthritis is a form of rheumatism in which the joints have become inflamed.
9.6.2 Arthritis
9.6.2.1 The term arthritis refers to many different diseases, the most common of which are rheumatoid arthritis (RA), Osteoarthritis, and gouty arthritis. All are characterized by inflammation of one or more joints. Inflammation, pain, and stiffness may also be present in adjacent parts of the body, such as the muscles near the joint.
9.6.3 Rheumatoid Arthritis (RA) (ROO-ma-toyd)
9.6.3.1 This is an autoimmune disease in which the body attacks its own tissues; in this case its own cartilage and joint linings. It is characterized by inflammation of the joint, swelling, pain, and loss of function. Usually, this form occurs bilaterally-if your left wrist is affected; your right wrist may also be affected, although usually not to the same degree.
9.6.3.2 Symptom
9.6.3.2.1 The primary symptom of rheumatoid arthritis is inflammation of the synovial membrane. If untreated, the following sequential pathology may occur. The membrane thickens and synovial fluid accumulates. The resulting pressure causes pain and tenderness. The membrane then produces an abnormal granulation tissue called a pannus, which adheres to the surface of the articular cartilage. The pannus formation sometimes erodes the cartilage completely. When the cartilage is destroyed fibrous tissue joins the exposed bone ends. The tissue ossifies and fuses the joint so that it is immovable- the ultimate crippling effect of rheumatoid arthritis. Most cases do not progress to this stage, but the range of motion of the joint is greatly inhibited by the server inflammation and w\swelling. The growth of the pannus is shat causes the distortion of the fingers that is so typical of the clinical appearance of hands that have been affected by rheumatoid arthritis. A plastic surgeon or orthopedic will remove the pannus growth before crippling and dysfunctional deformities have occurred-long before the joints are fused to the point of immobility.
9.6.3.3 Treatment
9.6.3.3.1 Treatment is aimed at reducing pain and inflammation and preserving muscle strength and joint function. Therapies include adequate rest; anti-inflammatory drugs, such as aspirin and in selected patients steroids; exercises to maintain full range of joint motion; heat and other forms of physical therapy; and weight loss to relieve pressure on weight-bearing joints. In severe cases, damaged joints may be surgically replaced, either partly or entirely, with artificial joints. The artificial parts are inserted after removal of the diseased portion of the articulating bone and its cartilage. The new metal or plastic joint is fixed in place with special acrylic cement. When freshly mixed in the operating room, it hardens as strong as bone in minutes.
9.6.3.4 Osteoarthritis (os’-tē-ō-ar-THRĪ-tis)
9.6.3.4.1 A degenerative joint disease far more than rheumatoid arthritis , and usually less damaging, is Osteoarthritis. It apparently results from a combination of aging, irritation of the joints and wear and abrasion.
9.6.3.4.2 Degenerative joint disease is a non-inflammatory, progressive disorder of movable joint, particularly weight-bearing joints. It is characterized by the deterioration of articular cartilage and by formation of new bone in the subchondral areas and at the margins of the joint. The cartilage slowly degenerates, and as the bone ends become exposed, small bumps, or spurs, of new osseous tissue are deposited on them. These spurs decrease the space of the joint cavity and restrict joint movement. Unlike rheumatoid arthritis, Osteoarthritis usually affects only the articular cartilage. The synovial membrane is rarely destroyed, and other tissues are unaffected. The main distinction between Osteoarthritis and rheumatoid arthritis is that the former strides the big joints (knees, hips) first, whereas the latter strikes the small joints first. But Osteoarthritis may affect the fingers, and when this is the case, the distal phalanges show the most prominent changes. The effect of rheumatoid arthritis on the fingers is most pronounced proximally, in the wrists and in the Metacarpophalangeal, and proximal Interphalangeal joints. Also, rheumatoid arthritis is more likely to be bilaterally symmetrical than is the case for Osteoarthritis.
9.6.3.5 Gouty Arthritis (GOW-tē)
9.6.3.5.1 Description
9.6.3.5.1.1 Uric acid (a substance that gives urine its name) is a waste product produced during the metabolism of nucleic acids. The person who suffers from gout either produces excessive amounts of uric acid or is not able to excrete normal amounts. The result is a buildup of uric acid in the blood. This excess acid then reacts with sodium to form a salt called sodium urate. Crystals of this salt are deposited in soft tissues. Typical sites are the kidneys, first metatarsal Phalangeal joint and the cartilage of the ears and other joints.
9.6.3.5.2 Cause
9.6.3.5.2.1 In gouty arthritis, sodium urate crystals are deposited in the soft tissues of the joints. The crystals irritate the cartilage, causing inflammation, swelling, and acute pain. Eventually, the crystals destroy all the joint tissues. If the disorder is not treated, the ends of the articulating bones fuse and the joint becomes immovable.
9.6.3.5.3 Treatment
9.6.3.5.3.1 Gouty arthritis occurs primarily in middle-aged and older males. It is believed to be the cause of 2 to 5 percent of all chronic joint diseases. Numerous studies indicate that an abnormal gene sometimes causes gouty arthritis. As a result of this gene, the body manufactures unusually large amounts of uric acid. Diet and environmental factors such as stress and climate are also suspected causes of gouty arthritis.
9.6.3.5.3.2 Although other forms of arthritis cannot be treated with complete success, the treatment of gouty arthritis with the use of various drugs has been quite effective. A chemical called colchicines has been utilized periodically since the sixth century to relieve the pain, swelling, and tissue destruction that occur during attacks of gouty arthritis. This chemical is derived from the variety of crocus plant from which the spice saffron is obtained. Other drugs, which either inhibit uric acid production or assist in the elimination of excess uric acid by the kidneys, are used to prevent further attacks. The drug allopurinal is used for the treatment of gouty arthritis because it prevents the formation of uric acid without interfering with nucleic acid synthesis. Rest and the use of anti-inflammatory medication may obtain relief from an attack of gouty arthritis.
9.6.4 Bursitis
Back Table of Contents References
9.6.4.1 An acute chronic inflammation of a bursa is called bursitis. The condition may be caused by trauma, by an acute or chronic infection (including syphilis and tuberculosis), or by rheumatoid arthritis. Repeated excessive friction often results in a bursitis with local inflammation and the accumulation fo fluid. Bunions are frequently associated with a friction bursitis over the head of the first metatarsal bone. Symptoms include pain, swelling, tenderness, and the limitation of motion involving the inflamed bursa. The prepatellar or subcutaneous Infrapatellar bursae may become inflamed in individuals who spend a great deal of time kneeling. This bursitis is usually called “housemaid’s knee” (“carpet layer’s knee”)
9.6.5 Dislocation
9.6.5.1 A dislocation, or luxation (luks-Ā-shun), is the displacement of a bone from a joint with tearing of ligaments, tendons, and articular capsules. A partial or incomplete dislocation is called a Subluxation. The most common dislocations are those involving a finger or shoulder. Those of the mandible, elbow, knee, or hip are less common. Symptoms include loss of motion, temporary paralysis of the involved joint, pain k, swelling, and occasionally shock. A dislocation is usually caused by a blow or fall., although unusual physical effort may lead to this condition.
9.6.6 Sprain and Strain
9.6.6.1 A sprain is the forcible wrenching or twisting of a joint with partial rupture or other injury to its attachments without luxation. It occurs when the attachments are stressed beyond their normal capacity. There may be damage to the associated blood vessels, muscles, tendons, ligaments, or nerves. A sprain is more serious than a strain, which is the overstretching of a muscle. Severe sprains may be so painful that the joint cannot be moved. There is considerable swelling and pain may occur owing to underlying hemorrhage from ruptured blood vessels. The ankle joint is most often sprained; the low back area is another frequent location for sprains.
10 Muscle Tissue
Back Table
of Contents References
10.1 Characteristics
10.2 Functions
10.3 Types
10.4 Skeletal Muscle Tissue
10.4.1
10.4.2 Connective Tissue Components
10.4.3 Nerve and Blood Supply
10.4.4 Histology
10.4.4.1 Myofibers (Muscle Fibers or Cells)
10.4.4.1.1 Skeletal muscles consist of thousands of elongated cylindrical cells called Myofibers or muscle fibers. The fibers lie parallel to one another and range from 10 to 100 micro meters (µm) in diameter. Some muscle fibers may reach lengths of 30 cm (12 inches) or more.
10.4.4.2 Sarcolemma (sarco=flesh; lemma=sheath)
10.4.4.2.1 Each muscle fiber is enveloped by a plasma membrane, which contains the cytoplasm of the muscle cell called the Sarcoplasm.
10.4.4.3 Sarcoplasm
10.4.4.3.1 The cytoplasm to the muscle cell that contains many nuclei (multinucleate) nestled close to the Sarcolemma. The Sarcoplasm also contains myofibrils (Discussed below), mitochondrion, and enzymes.
10.4.4.4 Sarcoplasmic reticulum (sar’-Kō-PLAZ-mik re-TIK-yoo-lum)
10.4.4.4.1 Membrane-enclosed tubules similar to smooth endoplasmic reticulum (A network of channels running through the cytoplasm) (Substance within plasma membrane) of other cells of the body which serves in intracellular transportation, support storage, synthesis and packaging of molecules.)
10.4.4.5 Terminal Cisterns
10.4.4.5.1 These are dilated sacs of sarcoplasmic reticulum, which form ring like channels around the myofibrils.
10.4.4.6 Transverse tubles (T tubules)
10.4.4.6.1 These run transversely through the fiber and perpendicularly to the sarcoplasmic reticulum and are extensions of the Sarcolemma that open to the outside of the fiber.
10.4.4.7 Triad
10.4.4.7.1 Consists of a transverse tubule and terminal cisterns on either side.
10.4.4.8 Myofibrils
10.4.4.8.1 Within the muscle cell there are 100-1000 cylindrical bundles of muscle fiber 1 or 2 micro-meters (µm) in diameter composed of thin and thick myofilaments.
10.4.4.9 Myofilaments
10.4.4.9.1 Thin Myofilaments
10.4.4.9.1.1 6 nm in diameter
10.4.4.9.2 Thick Myofilaments
10.4.4.9.2.1 16 nm in diameter
10.4.4.10 Sarcomeres
10.4.4.10.1 A compartment containing thin and thick Myofilaments comprising the contractile unit bounded on either end by Z lines.
10.4.4.11 Z Lines
10.4.4.11.1 Dense material on either end of the sarcomere, which define the contractile unit.
10.4.4.12 A (Anisotropic) Band
10.4.4.12.1 Represents the length of thick myofilaments, which are darkened at either end by overlapping with the thin myofilaments attached to the Z lines. The length of the darkening is increased with muscle contraction.
10.4.4.13 I (Isotropic) Band
10.4.4.13.1 Contains thin myofilaments only and has a Z line in the middle representing the thin myofilaments of two adjacent contractile units. The combination of alternating dark A bands and light I bands gives the muscle fiber its striated (striped) appearance.
10.4.4.14 H Zone
10.4.4.14.1 A region in the center of the A band containing thick myofilaments only.
10.4.4.15 M Line
10.4.4.15.1 The adjacent thick myofilaments are connected in the middle by these fine threads.
10.4.4.16 Actin
10.4.4.16.1 Thin myofilaments are anchored to the Z lines and project in both directions. Thin myofilaments are composed mostly of the protein actin. The actin molecules are arranged in two single strands that entwine helically and give the thin myofilaments their characteristic shape.
10.4.4.17 Myosin-Binding Site
10.4.4.17.1 The actin molecule has a myosin- binding site that interacts with a cross bridge of a myosin molecule (discussed below).
10.4.4.18 Tropomyosin and Troponin (Tropomyosin-troponin Complex)
10.4.4.18.1 Proteins which help regulate muscle contraction
10.4.4.18.2 Tropomyosin
10.4.4.18.2.1 Arranged in strands that are loosely attached to the actin helices.
10.4.4.18.3 Troponin
10.4.4.18.3.1 Located at regular intervals on the surface of Tropomyosin with three subunits
10.4.4.18.3.2 Troponin I
10.4.4.18.3.2.1 Actin binding
10.4.4.18.3.3 Troponin C
10.4.4.18.3.3.1 Calcium binding
10.4.4.18.3.4 Troponin T
10.4.4.18.3.4.1 Tropomyosin binding
10.4.4.19 Myosin
10.4.4.19.1 Thick myofilaments are composed mostly of the protein myosin, overlap the free ends of the thin myofilaments and occupy the A band of the sarcomere.
10.4.4.19.2 The myosin molecule is golf club shaped, with tails (Golf club handles) arranged parallel to one another, which form the shaft of the myofilament. The heads of the golf clubs project outward from the shaft and are arranged spirally of the surface of the shaft.
10.4.4.20 Cross Bridges, Actin-Binding Site and ATP-Binding Site
10.4.4.20.1 The projecting heads of the myosin molecule are referred to as cross bridges and contain an Actin-Binding Site and an ATP-Binding Site
10.5 Contraction
10.5.1 Sliding-Filament Theory
10.5.1.1 The myosin cross bridges pull on the thin myofilaments causing them to slide inward toward the H zone. The lengths of the thin and thick myofilaments do not change but the sarcomere shortens evidenced by the reduced space between the Z lines. The myosin cross bridges of the thick myofilaments connect with portions of actin of the thin myofilaments. The myosin cross bridges move like the oars of a boat on the surface of the tin myofilaments, and the thin and thick myofilaments slide past each other as the cross bridges pull on, the thin myofilaments. The sliding of myofilaments and shortening of sarcomeres causes the shortening of the muscle fibers.
10.5.2 Neuromuscular Junction
10.5.2.1 Neuron
10.5.2.1.1 Provides the stimulus for muscle contraction.
10.5.2.1.2 The thread-like process which delivers the nerve impulse is called an axon can be 91 cm (3 feet) or more in length before attaching to a muscle.
10.5.2.1.3 A bundle of axons from many different neurons compose a nerve.
10.5.2.2 Motor Neuron
10.5.2.2.1 This is a neuron, which only stimulates muscles.
10.5.2.3 Motor End Plate
10.5.2.3.1 Upon entering the skeletal muscle the axon of a motor neuron branches into axon terminals (telodendria) that come in contact with the Sarcolemma of individual muscle cells.
10.5.2.3.2 This region of Sarcolemma axon contact is known as the motor end plate.
10.5.2.4 Neuromuscular Junction (Myoneural junction (MNJ))
10.5.2.4.1 Refers to both the axon terminal of a motor neuron along with the motor end plate.
10.5.2.5 Synaptic end Bulbs
10.5.2.5.1 These are bulb like structures of the distal end of axon terminals at the neuromuscular junction.
10.5.2.6 Synaptic Vesicles
10.5.2.6.1 Membrane enclosed sacs that store neurotransmitters.
10.5.2.7 Neurotransmitters
10.5.2.7.1 These chemicals determine whether an impulse is passed on to a muscle (or gland or another nerve cell).
10.5.2.8 Synaptic Gutter (Trough)
10.5.2.8.1 This is the invaginated area of the Sarcolemma under the axon terminal.
10.5.2.9 Synaptic Cleft
10.5.2.9.1 The space between the axon terminal and Sarcolemma
10.5.2.10 Subneural Clefts
10.5.2.10.1 These are folds of the Sarcolemma along the synaptic gutter.
10.5.2.10.2 This greatly increases the surface area of the synaptic gutter allowing for increased numbers of receptor site molecules that are able to bond to the neurotransmitter.
10.5.2.11 Acetylcholine (as’-ē-til-KŌ-lēn) (Ach)
10.5.2.11.1 Released from the synaptic vesicles (and possibly the cytoplasm) when a nerve impulse (Action Potential) reaches the axon terminals and synaptic end bulbs at the neuromuscular junction.
10.5.2.11.2 After release Ach diffuses across the synaptic cleft and combines with receptor sites on the Sarcolemma of the muscle cell.
10.5.2.11.3 This combination alters the permeability of the Sarcolemma to sodium (Na+) and potassium (Ka+) ions, and ultimately results in the development of a muscle action potential that travels along the Sarcolemma and is responsible for initiating the muscle contraction.
10.5.2.11.4 For the vast majority of muscle cells there is only one neuromuscular junction for each cell located in the middle of the muscle cell.
10.5.2.11.5 The muscle action potential spreads from the center of the Sarcolemma of the cell to the ends and through the transverse tubules.
10.5.2.11.6 This permits nearly simultaneous contraction of all sarcomeres in the muscle cell as the action potential spreads deeply into the Sarcoplasm by way of the transverse tubules of the Sarcolemma.
10.5.3 Motor Unit
10.5.3.1 The motor unit includes the motor neuron and all the muscle fibers (cells) it stimulates.
10.5.3.2 150 muscle fibers may be stimulated by just one motor neuron depending on the region of the body.
10.5.3.3 When one neuron of a motor unit is stimulated this can result in the simultaneous contraction and relaxation of all 150 muscle fibers of a motor unit that receive sufficient stimulation.
10.5.3.4 When precise movements are needed such as for eye movements, these movements are carried out by the extrinsic (external) eye muscles and there are fewer than 10 muscle fibers to each motor unit.
10.5.3.5 Gross movements in muscles like the gastrocnemius and biceps brachii may have as many as 200 muscle fibers in each motor unit.
10.5.3.6 Recruitment (Motor Unit Summation)
10.5.3.6.1 Since stimulation of one motor neuron produces a contraction in all the muscle fibers in a particular motor unit the total tension in a muscle can be varied by adjusting the number of motor units that are activated.
10.5.3.6.2 The process of increasing the active number of motor units is called recruitment and is determined by the demands that activity places on the body.
10.5.3.6.3 Because not all motor units are active at the same time (Neurons fire asynchronously=Some muscles inhibited others excited) this prevents fatigue while maintaining contraction by allowing a brief rest for the inactive units.
10.5.3.6.4 Thus contractions can be maintained for long time periods with alternated smooth transitions between active and inhibited motor units.
10.5.3.6.5 Continuous muscle tone and a state of partial contraction as found in postural muscles
10.5.3.6.6 Recruitment helps produce smooth movements during muscle contraction rather than a series of jerky movements.
10.5.4 Physiology of Contraction
10.5.4.1 A relaxed muscle has a low concentration of calcium ions (Ca2 +) in the Sarcoplasm.
10.5.4.2 Calcium Ions are stored in the sarcoplasmic reticulum.
10.5.4.3 A relaxed muscle has a high concentration of ATP (Adenosine Triphosphate=(a-DEN-ō-sēn tri=FOS=fat)) and is attached to the ATP-binding sites of the myosin cross bridges.
10.5.4.4 The binding of ATP to the myosin cross bridges and the binding of the Tropomyosin-troponin complex to actin prevents the myosin cross bridges from combining with actin of the thin myofilaments.
10.5.4.5 When a nerve impulse (nerve action potential) reaches the synaptic end bulb of an axon terminal, a small amount of calcium enters the synaptic end bulb, causing the synaptic vesicles (and possibly the cytoplasm) to release Acetylcholine into the synaptic cleft.
10.5.4.6 It diffuses across the synaptic cleft and combines with receptor sites on the Sarcolemma of the muscle Fiber directly beneath the synaptic end bulb.
10.5.4.7 Acetylcholine-Gated Ion Channels
10.5.4.7.1 The combination of Acetylcholine with the receptor sites on the Sarcolemma of the muscle fiber causes a change in the permeability of the Sarcolemma in which membrane proteins called Acetylcholine-gated ion channels open and permit a rapid influx of positive ions, mainly Na+.
10.5.4.8 Muscle Action Potential
10.5.4.8.1 The muscle action potential travels along the Sarcolemma and then into the transverse tubules.
10.5.4.8.2 When the muscle action potentials conveyed over the transverse tubules close to the sarcoplasmic reticulum, the action potential causes the reticulum to release some of the calcium ions from storage into the Sarcoplasm surrounding the myofilaments.
10.5.4.9 Calcium Release Channels
10.5.4.9.1 These are voltage-sensitive, tunnel-shaped proteins in the sarcoplasmic reticulum which regulate the release of calcium from the sarcoplasmic reticulum.
10.5.4.10 Troponin (Calcium-binding protein)
10.5.4.10.1 Undergoes structural changes when a portion of it combines with calcium ions.
10.5.4.10.2 The Tropomyosin-troponin complex moves into a groove between actin strands, thus exposing the myosin-binding sites on actin.
10.5.4.11 . ATP (Adenosine Triphosphate=(a-DEN-ō-sēn tri=FOS=fat))
10.5.4.11.1 Muscle contraction requires energy, as well as calcium ions. The energy is supplied by ATP
10.5.4.11.2 ATP is attached to ATP-binding sites on myosin cross bridges.
10.5.4.11.3 Myosin cross bridges function as ATPase enzymes which can split ATP into ADP (adenosine diphosphate) + P with the release of energy.
10.5.4.11.4 The muscle action potential stimulates a muscle fiber and the myosin cross bridges functioning as ATPase enzymes, split ATP into ADP + P. The ADP + P remain attached to their binding sites on the myosin cross bridges, activated (energized) by the energy from the splitting of ATP and combine with myosin-binding sites on actin.
10.5.4.12 Power Stroke
10.5.4.12.1 Once the activated myosin cross bridges are attached to the myosin-binding sites on actin the activated myosin cross bridges change their orientation.
10.5.4.12.2 The myosin cross bridge moves toward the H zone in the center of the sarcomere and in so doing applies force to the thin actin myofilaments.
10.5.4.12.3 This movement of the myosin cross bridges is called the power stroke and causes the thin actin myofilaments to slide past the thick myosin myofilaments.
10.5.4.13 Detachment of Myosin Cross Bridge
10.5.4.13.1 Once the power stroke is complete, ATP combines with the ATP-binding sites on the myosin cross bridges, resulting in the detachment of the myosin cross bridge from actin.
10.5.4.14 Process Repeated
10.5.4.14.1 With the continued presence of a muscle action potential and Ca ions the process repeats itself e.g. ATP is split and a given myosin cross bridge then combines with another myosin-binding site further along the actin strand.
10.5.4.14.2 The myosin cross bridges keep moving back and forth like the cogs of a ratchet with each power stroke, moving the tin actin myofilaments toward the H zone.
10.5.4.14.3 This continual movement applies the force that causes the Z lines of a sarcomere to be drawn toward each other, and the sarcomere shortens.
10.5.4.14.4 The combined shortening of many sarcomeres results in global muscle contraction of the entire motor unit.
10.5.4.15 Maximal Muscle Contraction
10.5.4.15.1 During maximal muscle contractions, the distance between Z lines can be shortened to 50 % of the resting length.
10.5.4.16 Isometric Contraction
10.5.4.16.1 This cross bridge (contraction) does not always result in shortening of the muscle fibers and muscle.
10.5.4.16.2 When the contraction does not result in muscle shortening, it is called isometric contraction.
10.5.4.16.3 Using postural muscles to stand or sit is an example of an isometric contraction.
10.5.4.17 Acetylcholinesterase (AchE)
10.5.4.17.1 Acetylcholine is rapidly destroyed by an enzyme called Acetylcholinesterase (AchE). This allows the muscle fiber to go from a contracted state back to a relaxed state.
10.5.4.17.2 This enzyme is found on the surfaces o the subneural clefts of the Sarcolemma of muscle fibers.
10.5.4.17.3 As long as no further action potentials are generated there will be no new release of Ach and this stops the generation of a muscle action potential.
10.5.4.17.4
10.5.4.18 Calsequestrin and Calcium-ATPase
10.5.4.18.1 After the muscle action potential ends, the calcium ions are actively transported from the troponin of the thin filaments in the Sarcoplasm back into the sarcoplasmic reticulum for storage.
10.5.4.18.2 This is accomplished by two proteins called Calsequestrin and Calcium-ATPase and involves an expenditure of some ATP, as the molecule is broken down to release energy used to actively transport calcium ions into storage in the sarcoplasmic reticulum.
10.5.4.19 Tropomyosin-Troponin Complex
10.5.4.19.1 Once calcium ions are removed from the Sarcoplasm, the Tropomyosin-Troponin Complex is reattached to the actin strands, so that the myosin-binding sites of actin become covered and myosin cross bridges separate from actin and cannot reattach.
10.5.4.19.2 ATP is also required to detach the cross bridges.
10.5.4.20 Thin Myofilaments
10.5.4.20.1 Since the myosin cross bridges are broken, the thin myofilaments slip back to their relaxed position.
10.5.4.21 ADP
10.5.4.21.1 After the muscle action potential ends, ADP is resynthesized into ATP, which again attaches to the ATP-binding site of the myosin cross bridge.
10.5.4.21.2 The sarcomeres are thereby returned to their resting lengths, and the muscle fiber resumes its resting state.
10.5.4.21.3 During extensibility, sarcomere length can be increased by about 20 percent of the resting length.
10.5.4.22 Summary
10.5.4.22.1 Nerve Impulse (Action Potential)
10.5.4.22.1.1 The action potential of the motor nerve causes synaptic vesicles in motor axon synaptic end bulbs to release Acetylcholine (Ach).
10.5.4.22.2 Acetylcholine Diffusion and Muscle Action Potential
10.5.4.22.2.1 Acetylcholine diffuses across the synaptic cleft within the neuromuscular junction and initiates a muscle action potential that spreads over the surface of the Sarcolemma and transverse tubules.
10.5.4.22.3 Calcium Ionic Release
10.5.4.22.3.1 The muscle action potential enters the transverse tubules and sarcoplasmic reticulum and stimulates the sarcoplasmic reticulum to release calcium ions from storage into the Sarcoplasm.
10.5.4.22.4 Myosin-Binding Sites Exposed
10.5.4.22.4.1 Calcium ions combine with troponin, causing the Tropomyosin-troponin complex to move, thus exposing the myosin-binding sites on actin.
10.5.4.22.5 Power Stroke
10.5.4.22.5.1 When a muscle action potential stimulates a muscle fiber, ATPase splits ATP into ADP + P and energy is released.
10.5.4.22.5.2 The released energy activates (energizes) myosin cross bridges, which combine with the exposed myosin-binding sites on actin, and apply force which causes the myosin cross bridges to move toward the H zone (Power Stroke).
10.5.4.22.5.3 This movement results in the sliding of the thin myofilaments past thick myofilaments.
10.5.4.22.6 Z Lines Narrow
10.5.4.22.6.1 The sliding draws the Z lines toward each other, the sarcomere shortens, the muscle fibers contract and the muscle contracts.
10.5.4.22.7 Acetylcholine Inactivated
10.5.4.22.7.1 Acetylcholine is inactivated by Acetylcholinesterase (AchE). As a result, Ach no longer has any effect at the neuromuscular junction.
10.5.4.22.8 Calcium Ions Reabsorbed
10.5.4.22.8.1 In the absence of further action potentials, calcium ions are actively transported back into the sarcoplasmic reticulum by Calsequestrin and calcium ATPase, using energy from ATP break down.
10.5.4.22.9 Tropomyosin-troponin complex Reattached
10.5.4.22.9.1 The low calcium concentration in the Sarcoplasm permits the Tropomyosin-troponin complex to reattach to actin.
10.5.4.22.9.2 As a result, myosin-binding sites of actin become covered, myosin cross bridges separate from actin.
10.5.4.22.9.3 ADP is resynthesized into ATP (which reattaches to the ATP-binding site of the myosin cross bridge), and the thin myofilaments return to their relaxed position.
10.5.4.22.10 Sarcomeres at Resting Length
10.5.4.22.10.1 Sarcomeres return to their resting lengths, muscle fibers relax, and the muscle relaxes.
10.5.4.23 Clinical Application
10.5.4.23.1 Rigor Mortis
10.5.4.23.1.1 Following death, because of a lack of ATP, myosin cross bridges remain attached to the actin myofilaments, thus preventing relaxation.
10.5.4.23.1.2 The myosin cross bridges need a constant stream of ATP to both attach and detach to the actin myofilaments. Thus the lack of ATP makes detachment impossible
10.5.4.23.1.3 The resulting condition in which muscles are in a state of rigidity (cannot contract or stretch) is called rigor mortis (rigidity of death).
10.5.4.23.1.4 The time elapsing between death and the onset of rigor mortis varies greatly among individuals.
10.5.4.23.1.5 Those who have had long, wasting illnesses undergo rigor mortis more quickly.
10.5.4.23.1.6 Rigor mortis is not a permanent state that continues unabated after death.
10.5.4.23.1.7 Depending on conditions, it lasts about 24 hours, then begins to abate, and disappears after another 12 hours when tissues begin to disintegrate, although a residual stiffness in the joints will remain.
10.5.4.23.2 McArdle's Disease McArdle's Disease
10.5.4.23.2.1 Is caused by a defect in an enzyme, which breaks down glycogen into glucose.
10.5.4.23.2.2 Since glycogen is stored in muscle tissue as a reserve energy supply for muscle contraction it cannot be used for that purpose if it isn’t converted to glucose by a defective enzyme.
10.5.4.23.2.3 The symptoms of this disease include, severe cramping and or pain during exercise and muscle weakness later in life.
10.5.4.23.3 Carnatine Deficiency Carnatine Deficiency
10.5.4.23.3.1 It serves to transport fatty acids across the cell membrane and into the mitochondria, where these fatty acids are burned for energy. It increases the rate of oxidation of fats in the liver, and this suggests that it plays a role in improving energy generation from this angle as well.
10.5.5 Energy for Contraction and Relaxation
10.5.5.1 Phosphagen System
10.5.5.1.1 ATP
10.5.5.1.1.1 ATP is the immediate source of energy for muscle contraction and is broken down by ATPase into ADP + P and energy synthesized by muscle fibers as follows; ADP + P+ Energy=ATP.
10.5.5.1.1.2 The amount of ATP present in the muscle fibers is sufficient to maintain muscle contraction during vigorous exercise for 5 to 6 seconds.
10.5.5.1.2 Phosphocreatine (fos’fō-KRĒ-a-tin)
10.5.5.1.2.1 Skeletal muscle fibers contain this high-energy molecule, which is used to generate ATP rapidly.
10.5.5.1.2.2 There are about 2 to three times greater amounts of Phosphocreatine than ATP in the muscle fiber.
10.5.5.1.2.3 Phosphocreatine breaks down into creatine and phosphate and produces large amounts of energy as follows; Phosphocreatine=Creatine + Phosphate + Energy.
10.5.5.1.2.4 Some of the released energy is used to convert ADP to ATP and takes place in a fraction of a second to provide enough energy for muscles to contract maximally for about 15 seconds.
10.5.5.1.2.5 This energy system is used for maximal short bursts of energy and allows time for the rate of two metabolic processes, glycolysis and cellular respiration to be increased.
10.5.5.2 Glycogen-Lactic Acid System
10.5.5.2.1 Glycolysis
10.5.5.2.1.1 After Phosphocreatine depleted, then the source of energy is glucose, which is derived from the breakdown of glycogen (stored glucose), which is stored in the muscles, liver and found in the blood.
10.5.5.2.1.2 Glycogen breakdown is accomplished with calcium ions and a calcium-binding protein called calmodulin.
10.5.5.2.1.3 Once glucose is produced by this breakdown, each molecule of glucose is split into two molecules of pyruvic acid, a process called glycolysis.
10.5.5.2.1.4 This process releases energy which is used to form ATP and because glycolysis does not require oxygen it is referred to as an anaerobic process summarized as follows; 1 Glucose=2 Pyruvic acid + Energy (ATP).
10.5.5.2.2 Glycogen-Lactic Acid System
10.5.5.2.2.1 When there is not sufficient oxygen for the complete catabolism of pyruvic acid, it is converted to lactic acid, some of which diffuses out of the muscle fibers and eventually into blood.
10.5.5.2.2.2 The production of lactic acid in this way releases energy from glucose that can be used to produce ATP, and it occurs anaerobically providing sufficient energy for about 30-40 seconds fo maximal muscle activity.
10.5.5.3 Aerobic System
10.5.5.3.1 Cellular Respiration
10.5.5.3.1.1 When sufficient oxygen is present, the pyruvic acid formed by glycolysis enters the mitochondria of muscle fibers, where it is completely catabolized to carbon dioxide and water in the metabolic process called cellular respiration.
10.5.5.3.1.2 This is considered an aerobic process since it requires oxygen.
10.5.5.3.1.3 The complete catabolism of pyruvic acid also yields energy that is used to generate most of a muscle fiber’s ATP: Pyruvic Acid + O2 =CO2 + H2O + Energy (ATP).
10.5.5.3.1.4 This process will continue as long as nutrients (fatty acids (from Fats), amino acids (from proteins), and glucose derived from glycogen breakdown or delivered to the muscle by way of blood) and adequate oxygen last.
10.5.5.4 Summary
10.5.5.4.1 Phosphagen System
10.5.5.4.1.1 15 seconds muscular activity=100-meter dash
10.5.5.4.2 Glycogen-lactic Acid System
10.5.5.4.2.1 30-40 seconds muscular activity=400-meter dash
10.5.5.4.3 Aerobic System
10.5.5.4.3.1 Prolonged activity as long as sufficient oxygen and nutrients are available=Jogging
10.5.6 All-or-None Principle
10.5.7 Muscle Tension
10.5.8 Muscle Tone
10.5.9 Muscular Atrophy and Hypertrophy
10.5.10 Types of Skeletal Muscle Fibers
10.6 Cardiac Muscle Tissue
10.7 Smooth Muscle Tissue
10.8 Regeneration of Muscle Tissue
10.9 Homeostasis
10.10 Aging and Muscle Tissue
10.11 Developmental Anatomy of the Muscular System
10.11.1
11
Muscular
System*
Back Table of Contents References
Link To Muscular System Chapter
12 Nervous Tissue*
12.1 Introduction
12.1.1
12.2 Organization
12.2.1
12.3 Histology
12.3.1
12.3.2 Neuroglia
12.3.2.1
12.3.3 Neurons
12.3.3.1
12.3.3.2 Structure
12.3.3.2.1
12.3.3.3 Structural Variation
12.3.3.3.1
12.3.3.4 Classification
12.3.3.4.1
12.4 Physiology
12.4.1
12.4.2 Nerve Impulse
12.4.2.1
12.4.2.2 Membrane Potentials
12.4.2.2.1
12.4.2.3 Excitability
12.4.2.3.1
12.4.2.4 All-or-None Principle
12.4.2.4.1
12.4.2.5 Saltatory Conduction
12.4.2.5.1
12.4.2.6 Speed of Nerve Impulses
12.4.2.6.1
12.4.3 Conduction across Synapses
12.4.3.1
12.4.3.2 Excitatory Transmission
12.4.3.2.1
12.4.3.3 Inhibitory Transmission
12.4.3.3.1
12.4.3.4 Integration at Synapses
12.4.3.4.1
12.4.4 Neurotransmitters
12.4.4.1
12.4.5 Regeneration
12.4.5.1
12.5 Organization of Neurons
12.5.1
12.6 Neurons and Supporting Cells Human Physiology Fox 150-151
12.6.1 Neurons produce and conduct electrochemical impulses and along with supporting cells comprise the nervous system.[1]
12.6.2 The nervous system is composed of neurons, which produce and conduct electrochemical impulses, and supporting cells, which assist the functions of neurons. Neurons are classified functionally and structurally; the various types of supporting cells perform specialized functions.
12.6.3
The nervous system is divided into the central nervous system (CNS),
which includes the brain and spinal cord, and the peripheral nervous system
(PNS), which includes the cranial nerves arising from the brain and the spinal
nerves arising from the spinal cord.
12.6.4
The nervous system is composed of only two principal types of cells-neurons
and supporting cells. Neurons are the basic structural and functional units of
the nervous system. They are specialized to respond to physical and chemical
stimuli, conduct electrochemical impulses, and release chemical regulators.
Through these activities, neurons enable the perception of sensory stimuli,
learning, memory, and the control of muscles and glands. Most neurons cannot
divide by mitosis, although many can regenerate a severed portion or sprout
small new branches under certain conditions.
12.6.5
Supporting cells aid the functions of neurons and are about five times
more abundant than neurons. In the CNS, supporting cells are collectively
called neuroglia, or simply glial cells (glia = glue). Unlike neurons, glial
cells retain limited mitotic abilities (brain tumors that occur in adults are
usually composed of glial cells rather than neurons).
12.6.6
Neurons
12.6.6.1 Although neurons vary
considerably in size and shape, they generally have three principal regions:
(1) a cell body, (2) dendrites, and (3) an axon (Illustration # 2). Dendrites and axons can
be referred to generically as processes or
extensions from the Cell body (Neuron).
12.6.6.2 The cell body, or perikaryon (peri= around; karyon = nucleus), is the enlarged portion of the neuron that contains the nucleus. It is the "nutritional center" of the neuron where macromolecules are produced. The cell body also contains densely staining areas of rough endoplasmic reticulum known as Nissl bodies that are not found in the dendrites or axon. The cell bodies within the CNS are frequently clustered into groups called nuclei (not to be confused with the nucleus of a cell). Cell bodies in the PNS usually occur in clusters called ganglia (Illustration # 2).
12.6.6.3
Dendrites
12.6.6.3.1 Dendrites (dendron = tree branch) are thin, branched processes that extend from the cytoplasm of the cell body. Dendrites provide a receptive area that transmits electrical impulses to the cell body.
12.6.6.4
Axon
12.6.6.4.1
The
axon is a longer process that conducts impulses away from the cell body. Axons
vary in length from only a millimeter long to up to a meter or more (for those
that extend from the CNS to the foot). The origin of the 'axon near the cell
body is an expanded region called the axon hillock; it is here that nerve impulses originate. Side branches
called axon collaterals may extend from the axon.
12.6.6.5 Proteins and other molecules are transported through the axon at faster rates than could be achieved by simple diffusion. This rapid movement is produced by two different mechanisms: axoplasmic flowAxoplasmic flow and axonal transport (table 7.2).
12.6.6.6
Axoplasmic flow
12.6.6.6.1 Axoplasmic flow, the slower of the two, results from rhythmic waves of contraction that push the cytoplasm from the axon hillock to the nerve endings. Axonal transport, which employs rnicrotubules and is more rapid and more selective, may occur in a reverse (retrograde) direction as well as in a forward (orthograde) direction. Indeed, retrograde transport may be responsible for the movement of herpes virus, rabies virus, and tetanus toxin from the nerve terminals into cell bodies.
12.6.7
Test Your
Knowledge Section Test
13 Spinal Cord and Spinal Nerves*
13.1
13.2
Grouping of Neural Tissue Principles of
A&P 355
13.2.1 Introduction
13.2.1.1 White matter consists of myelinated nerve fibers such as dendrites or axons, which are grouped together in tracks and supported by glial cells. Gray matter consists of unmyelinated axons or dendrites and nerve cell bodies. The absence of myelin in these areas accounts for their gray color. A tract is a bundle of nerve fibers consisting of dendrites and axons that run long distances up and down the spinal cord and connect parts of the brain. For our purposes, these tracks in the spinal cord consist of ascending sensory fibers or descending motor fibers. The spinal cord itself consists of both gray and white matter. The horns of the spinal cord consist of gray matter in the form of an H shape. If the H is situated with the top toward the anterior portion of the body the top of H shape is called the anterior Gray Horn and the bottom of the H shape is called the posterior Gray Horn. On the outside of the H is the lateral Gray Horn. The crossbar of the H is situated in the center of the spinal cord. The white columns are divided into anterior and posterior white columns and lateral white columns. Down the center of the spinal cord is a division much as in the brain and is called a sulcus. The two halves to the spinal cord are connected in the center, which is also, where the center of the H is situated. The crossbar of the H is known, as the gray commissure. At the center of the gray commissure is the central canal, which is contiguous with the fourth ventricle of the brain. Anterior to the gray commissure is the anterior white commissure which connects the two half’s of the spinal cord.
13.3
Spinal
Cord
13.3.1
Protection
and Coverings
13.3.1.1
Vertebral
Canal
13.3.1.1.1
13.3.1.2 Meninges Principles of A&P 355
13.3.1.2.1 The spinal cord successive layers of covering (meninges) with the outermost layer are known as the dura mater (tough mother). The meninges cover and protect the spinal cord and run continuously around the spinal cord and brain. Between the dura mater and the vertebral canal is the epidural space, which is filled with fat, connective tissue, and blood vessels. This tissue helps pad the spinal cord from injury. It is in the epidural space at the level of the second lumbar vertebrae that doctors will inject an anesthetic for both the pain for an irritated nerve or as a saddle block for childbirth. The second innermost covering of the spinal cord is the arachnoid (spider like). This web like structure is also continuous with the arachnoid of the brain. The subdural space is in between the arachnoid and the dura mater. The innermost covering of the spinal cord is called the pia mater (delicate mother) it connects with the spinal cord and although it is fibrous, it is transparent. It contains numerous blood vessels and nerves. The space between the arachnoid and the pia mater is the subarachnoid space. This space contains the cerebral spinal fluid. Inflammation of the meninges is known as meningitis. The most common form of meningitis is inflammation of the arachnoid and pia mater and is known as leptomeningitis.
13.3.2 General Features Principles of A&P 355
13.3.2.1 The spinal cord is about 1 1/2 ft. long and begins as a continuation of the medulla oblongata, which is the inferior part of the brain's stem, and extends from the foramen magnum of the occipital bone to the upper border of the 2nd lumbar vertebrae. It is cylindrical in shape and slightly flattened from the anterior to the posterior. The spinal cord is about 1 in. in circumference in the mid thoracic region but is enlarged in both the cervical (cervical enlargement) and the lumbar region (lumbar enlargement). The spinal cord forms an unbroken tube from its exit at the foramen magnum and the cranial cavity to the filum terminale and its connection at the coccyx. Its protective coverings are called meninges. The meninges of the spinal cord are continuous with the meninges of the brain. The filum terminale is a non-nervous fibrous tissue composed mostly of pia mater. It attaches to the end of the spinal cord at the conus medullaris, which is located at the intervertebral disc between the first and 2nd lumbar vertebrae. The filum terminale then attaches to the coccyx. The spinal cord is then securely anchored at two places, at both the cranium and the coccyx. The spinal cord is also attached along its entire route to the vertebral column, by denticulate ligaments, which act as shock absorbers in protecting the cord from injury.
13.3.3
Structure
in Cross Section
13.3.3.1
13.3.4
Functions
13.3.4.1
13.3.4.2
Impulse
Conduction
13.3.4.2.1
13.3.4.3
Reflex
Center
13.3.4.3.1
The second
principal function of the spinal cord is to serve as a center for some reflex
actions. Spinal nerves are the paths of
communication between his spinal cord tracts and the periphery. Each pair of spinal nerves is connected to a
segment of the cord by two points of attachment called roots. The posterior or dorsal (sensory) root
contains sensory nerve fibers only and conducts nerve impulses from the
periphery to the spinal cord. These
fibers extend into the posterior (dorsal) Gray horn. Each dorsal root also has a swelling, the posterior or dorsal
(sensory) root ganglion that contains the cell bodies of the sensory neurons
from the periphery. The other point of
attachment of a spinal nerve to the cord is the anterior or ventral (motor)
root. It contains motor neuron axons
only and conducts impulses from spinal cord to the periphery.
13.3.4.3.2
The cell
bodies of the motor neurons are located in the gray matter of the cord. If the motor impulse supplies the skeletal
muscle, the cell bodies are located in the anterior (ventral) Gray horn. If, however, the motor nerve impulse
supplies smooth muscle, cardiac muscle, or a gland through the autonomic
nervous system the cell bodies are located in the lateral gray horn.
13.3.4.4
Reflex
Arc and Homeostasis
13.3.4.4.1
Introduction
13.3.4.4.1.1 The path a nerve impulse follows from its origin in the dendrites or cell body of a neuron in one part of the body to its termination elsewhere in the body is called a conduction pathway. All conduction pathways consist of circuits of neurons. One pathway is known as a reflex arc, the functional unit of the nervous system. A reflex arc contains two or more types of neurons over which nerve impulses are conducted from a receptor to the brain or spinal cord by way of sensory neurons and then to an effector by way of motor neurons. The basic components of a reflex arc are as follows;
13.3.4.4.1.1.1 Receptor
13.3.4.4.1.1.1.1 The distal end of a dendrite or a sensory structure associated with the distal end of a dendrite. Its role in the reflex arc is to respond to a specific change in the internal or external environment by initiating a nerve impulse in a sensory neuron by way of a receptor potential (a local depolarization of the receptor cell membrane).
13.3.4.4.1.1.2 Sensory neurons
13.3.4.4.1.1.2.1 Passes the nerve impulse from the receptor to its axonal termination in the central nervous system.
13.3.4.4.1.1.3 Center
13.3.4.4.1.1.3.1 A region in the central nervous system where an incoming sensory impulse generates an outgoing motor impulse. In the center, the impulse may be inhibited, transmitted, or rerouted. In the center of some reflex arcs, the sensory neuron directly generates the impulse in the motor neuron. The center usually contains one or more association neurons between the sensory neuron and the motor neuron leading to a muscle or gland.
13.3.4.4.1.1.4 Motor neuron
13.3.4.4.1.1.4.1 Transmits the impulse generated by the sensory or association neuron in the center to the effector organ of the body that will respond, such as a muscle or gland.
13.3.4.4.1.1.5 Effector
13.3.4.4.1.1.5.1 The organ of the body that responds to the motor nerve impulse. This response is called a reflex and involves either an increase or decrease in muscle contraction or an increase or decrease in secretion by glands.
13.3.4.4.2 Reflexes
13.3.4.4.2.1 These are fast responses to certain changes (stimuli) in the internal or external environment that allow the body to maintain homeostasis. Reflexes are associated not only was skeletal muscle contraction but also with body functions such as heart rate, respiration, digestion, urination, and defecation, which involve cardiac and smooth muscle and glands. Reflexes carried out by the spinal cord alone are called spinal reflexes. Reflexes that result in contraction of skeletal muscles are known as somatic reflexes. Reflexes that involve brain centers cranial nerves are called cranial reflexes. Those that cause the contraction of smooth are cardiac muscle or secretion by glands are visceral (autonomic) reflexes. Our concern at this point is to examine a few somatic spinal reflexes: the stretch reflex, tendon reflex, flexor reflex and crossed extensor reflex.
13.3.4.4.3 Physiology of the Stretch Reflex
13.3.4.4.3.1 The stretch (tendon jerk) reflex is based on a two neuron or monosynaptic reflex arc. Only two types of neurons (one sensory and one motor) are involved, and there's only one synapse in the pathway. This reflex results in the contraction of a muscle when it is stretched suddenly. Slight stretching of the muscle stimulates receptors in the muscle called muscle spindles. The spindles monitor changes in the length of the muscle by responding to the rate and degree of change in length. Once the spindle is stimulated (depolarized to threshold), a nerve impulse is sent along a sensory neuron through the posterior root of the spinal nerves to the spinal cord. The sensory neuron synapses with a motor neuron in the anterior gray horn. The sensory neuron generates an impulse at the synapse that is transmitted along the motor neuron. The motor neuron in the anterior root of the spinal nerve terminates in a skeletal muscle. Once the impulse reaches the stretched muscle, a muscle action potential is generated and the muscle contracts. Thus, the stretch is counteracted by contraction, which shortens the muscle that had been stretched.
13.3.4.4.3.2 Since the sensory nerve impulse enters the spinal cord on the same side that the motor nerve impulse leaves the spinal cord, the reflex arc is called an ipsilateral (ip’si-LAT-er-al) reflex arc. All monosynaptic reflex arcs are ipsilateral as are deep tendon reflexes (discussed shortly).
13.3.4.4.3.3 The stretch reflex is essential in maintaining muscle tone and is important for muscle functions during exercise. It also helps prevent injury from overstretching of muscles. Moreover, it is the basis for several tests used in neurological examinations. One such reflex is the patellar reflex (knee jerk). This reflex involves extension of the leg by contraction of the quadriceps femoris muscle in response to tapping the patellar ligament. When the patellar ligament below the patella is tapped (stimulus), receptors (muscle spindles) in the quadriceps femoris muscle sensitive to changes in muscle length are stimulated because tapping the tendon of the muscle causes a rapid lengthening of the muscle. Sensory impulses from the receptors are sent to the spinal cord, and the returning motor nerve impulse results in generation of a muscle action potential that causes contraction of the muscle and extension of the leg at the knee, or a knee jerk as the muscle shortens during contraction.
13.3.4.4.3.4 Although the stretch reflex pathway contains only two types of neurons, other neurons are involved in the pathway in various ways. For example, the sensory neuron from the muscle spindle that enters the spinal cord also synapses with an inhibitory association neuron that, in turn, synapses with a motor neuron that controls antagonistic muscles. The inhibitory association neuron inhibits the motor neurons that normally send excitatory impulses to antagonistic muscles. Thus, during the stretch reflex when the stretched muscle is counteracted by contraction, antagonistic muscles that oppose the contraction are inhibited. This phenomenon, by which action potentials stimulate contraction of one muscle and simultaneously inhibit contraction of antagonistic muscles, is called reciprocal innervation. It avoids conflict between prime movers and antagonists and is vital in coordinating body movements. Since skeletal muscles act in groups rather than alone and that each muscle in the group (agonist, antagonist, synergist, fixator) has a specific role in bringing about the movement.
13.3.4.4.3.5 The sensory neuron from the muscle spindle also synapses with neurons that relay impulses to the brain by way of sensory tracts. In this way, the brain is provided with information about the state of stretch or contraction of skeletal muscles that enables it to coordinate muscular movements and posture.
13.3.4.4.4 Physiology of the Tendon Reflex
13.3.4.4.4.1 Reflexes other than the stretch reflex involve association neurons in addition to sensory and motor neurons. Since more than two types of neurons are involved, there is more than one type of synapse, and thus these reflexes are polysynaptic reflex arcs. One example of a reflex based on a polysynaptic reflex arc is the tendon reflex. The tendon reflex, like the stretch reflex, is also ipsilateral.
13.3.4.4.4.2 Just as the stretch reflex operates as a feedback mechanism to control muscle length, the tendon reflex operates as a feedback mechanism to control muscle tension by protecting tendons and their associated muscles from excessive tension. The receptors for this reflex are called tendon organs or Golgi tendon organs. Tendon organs lie within muscle tendons near the junction of a tendon and a muscle. Whereas muscle spindles are sensitive to changes in muscle length, tendon organs detect and respond to changes in muscle tension caused by passive stretch or muscular contraction.
13.3.4.4.4.3 When an increase in tension is applied to a tendon, the tendon organ is stimulated (depolarized to threshold) and nerve impulses are generated and transmitted to the spinal cord via a sensory neuron. Within the spinal cord, the sensory neuron synapses with an inhibitory association neuron, which, in turn, synapses with and hyperpolarizes (inhibits) a motor neuron that innervates the muscle associated with the tendon organ. Thus, as tension on the tendon organ increases, the frequency of inhibitory impulses increases, and the inhibition of the motor neurons to the muscle developing excess tension causes relaxation of the muscle. In this way, the tendon reflex protects the tendon and muscle from damage from excessive tension. Thus, the tendon reflex is protective.
13.3.4.4.4.4 The sensory neuron from the tendon organ also synapses with a stimulatory association neuron in the spinal cord. The stimulatory association neuron, in turn, synapses with motor neurons controlling antagonistic muscles. Thus, whereas the tendon reflex brings about relaxation of the muscle containing the tendon organ, it also brings about contraction of the antagonistic muscles. This is another example of reciprocal innervation. The sensory neuron also synapses with neurons that relay nerve impulses to the brain by way of sensory tracts, thus informing the brain about the state of muscle tension throughout the body.
13.3.4.4.5 Physiology of the Flexor Reflex and Crossed Extensor Reflex
13.3.4.5
Reflexes
and Diagnosis
13.3.4.5.1
13.4
Spinal
Nerves
13.4.1
Names
13.4.1.1
13.4.2
Composition
and Coverings
13.4.2.1
13.4.3
Distribution
13.4.3.1
Branches
13.4.3.1.1
13.4.3.2
Plexuses
13.4.3.2.1
13.4.3.2.2 Cervical Plexus
13.4.3.2.3 Brachial Plexus
13.4.3.2.4 Lumbar Plexus
13.4.3.2.5 Sacral Plexus
13.4.3.2.5.1 Sacral Plexus Nerves
13.4.3.2.5.1.1 Sciatic
13.4.3.2.5.1.1.1 Its nerve roots are L4-S3.. Composed of two nerves: tibial and common peroneal, bound together by common sheath of connective tissue.
13.4.3.2.5.1.1.2 It splits into its two divisions, usually at knee.
13.4.3.2.5.1.1.3 As the sciatic nerve descends through thigh, it sends branches to hamstring muscles (biceps femoris, semitendinousus, semimembranosus) and adductor magnus.
1.1.1.1.1.1.1.1.1 Tibial (medial popliteal)
1.1.1.1.1.1.1.1.2 Its nerve roots are L4-S3. Innervates gastrocnemius, Plantaris, Soleus, Popliteus, Tibialis posterior, flexor digitorum longus and flexor hallucis longus muscles.
1.1.1.1.1.1.1.1.3 Branches of tibial nerve in foot are medial plantar nerve and lateral plantar nerve.
1.1.1.1.1.1.1.1.4 Medial Plantar
1.1.1.1.1.1.1.1.5 Abductor hallucis, flexor digitorum brevis, and flexor hallucis brevis muscles, skin over medial two-thirds of plantar surface of foot.
1.1.1.1.1.1.1.1.6 Lateral Plantar
1.1.1.1.1.1.1.1.7 Remaining muscles of foot not supplied by medial plantar nerve; skin over lateral third of plantar surface of foot.
1.1.1.1.1.1.1.1.8 Common Peroneal (lateral popliteal)
1.1.1.1.1.1.1.1.9 Superficial Peroneal
1.1.1.1.1.1.1.1.10 Peroneus longus and Peroneus brevis muscles; skin over distal third of anterior aspect of leg and dorsum of foot.
1.1.1.1.1.1.1.1.11 Deep Peroneal
1.1.1.1.1.1.1.1.12 Tibialis anterior, extensor hallucis longus, Peroneus Tertius, and extensor digitorum longus and brevis muscles; skin over great and second toes.
13.4.3.3
Intercostal
(Thoracic) Nerves
13.4.3.3.1
13.4.4
Dermatomes
13.4.4.1
13.4.4.2 Dermal Segmentation Ciba 55
13.4.4.2.1
Sensations
from the outside world are carried to the consciousness by the spinal and some
cranial nerves to the brain. They travel over the posterior roots; their
further detailed course is for the moment " not of interest. All the cell
bodies are situated in ganglia outside of the central nervous system proper;
they have a peripheral branch going to the "sense organ and a central
branch going to the spinal cord or the brain.
13.4.4.2.2
We
include here the fibers carrying hot and cold, pain and touch. Vibration sense
and position sense are probably complicated sensations involving several of the
primitive sensations, including the proprioceptive sensations, which in the
spinal cord go also into the ganglia, but in the head region end in the
mesencephalic nucleus of the trigeminal nerve.
13.4.4.2.3
In the
head, sensations are mediated by the trigeminus and in a small region behind
the external auditory meatus by the facial and the vagus nerves. In the body,
the various spinal nerves take over. These are largely arranged metamerically,
that is to say, one segment lies behind the other in an orderly sequence.
However, these sequences are interrupted in the arm and the leg regions where
several segments have been carried out to the periphery of the limb and are no
longer represented near the axis of the body. Hence, on the ventral side of the
body, C5 borders on TI, and LI borders next to S2.
13.4.4.2.5
Two
other important points should be realized: The segmentations given in almost
all diagrams are based on pain, but the borders for the reception of pain are
not the same as those for the reception of, say, touch, which generally
involves a somewhat narrower sphere. The second point is that the different
nerves overlap to some extent, so that frequently no more than a hypesthesia
develops after one root has been cut.
13.4.4.2.6
If one
is familiar with the cutaneous distribution of various nerve roots; it is
possible to localize the site and level of any pathologic disturbance.
A chart outlining the exact sensory dermal segments serves as a good reference, but it is valuable to remember some surface landmarks which will serve as a general guide to localization: Illustration #4 Illustration #4b
13.4.4.2.8
Cervical
five sensory roots supply the clavicular region.
13.4.4.2.9
Cervical
five and six sensory roots supply the deltoid region.
13.4.4.2.10
Thoracic
four sensory roots supply the nipple area.
13.4.4.2.11
Thoracic
10 sensory roots supply the umbilicus.
13.4.4.2.12
The
groin region is supplied by thoracic I2 sensory root. .
13.4.4.2.13
Cervical
5,6 and 7 sensory roots supply the lateral aspect of the arm.
13.4.4.2.14
The
inner aspect of the arm is supplied by cervical eight and thoracic one sensory
roots.
13.4.4.2.15
Lumbar
L1, 2, 3 and 4 sensory roots supply the inner and anterior surfaces of the
thigh from above down.
13.4.4.2.16
Lumbar
5 and sacral I and 2 sensory roots supply the outer and posterior surfaces of
the thigh. "
13.4.4.2.17
Sacral
2, 3, 4 and 5 sensory roots supply the perineum.
13.4.4.2.18
The
hand is supplied from the radial to the ulnar borders by cervical 6, 7 and 8
sensory roots. Cervical 6 supplies the thumb, cervical 7 the middle of the
hand, including the index and middle fingers, and cervical 8 the ulnar border,
including the ring and little fingers.
13.4.4.2.19
Sacral
1, lumbar 5 and lumbar 4 sensory roots supply the foot from its lateral to its
medial surface.
13.4.4.2.20 NOTE: Some discrepancies exist in the definition of the dermal segments as published by different authors, but the chart produced here is based essentially on the work of Dr. J. Jay Keegan, University of Nebraska C[2]ollege of Medicine. We herewith express our appreciation for his contributions and personal communication. [3] [4] [5]
13.4.4.3 Dermatome??
13.4.4.3.1 It is something of a miracle, that we evolve into what we are today from such humble beginnings. From just a few cells in the womb of our mother, we become what we are today. The Fetal stem cells form the organs, bones, muscles, nerves and fascia. The specialized set of cells that form the fascia including the skin is called a dermatome. The word dermatome has also come to mean the area of the skin supplied by the dorsal RAMI of the spinal nerves. Each dermatome is named after a spinal level from which the nerve originates. For example, the L 1 dermatome is supplied by the first lumbar nerve. The L5 dermatome is supplied by the 5th lumbar nerve. These sensory nerves allow us to feel both the caressing touch of a loved one, and the burn of a frying pan.
13.4.4.3.2
Our
muscles, bones & skin form when we are still in the womb.
The dermatome is the name for a specialized set of cells in the developing fetus, which forms the connective tissues, including the dermis or skin. Illustration # 4
13.4.4.3.4
–The skin over
the entire body is supplied segmentally by spinal nerves.
13.4.4.3.5 The skin segment supplied by the dorsal root of a spinal nerve is also called a dermatome.
13.4.5
Arteries of the Spinal Cord Ciba 53 Illustration # 5
13.4.5.1 The spinal cord derives
its blood supply from the vertebral artery and from a series of spinal rami,
which enter the intervertebral foramina at successive levels.
13.4.5.2 The posterior spinal
artery is a branch of the vertebral artery, which begins near the lateral
margin of the medulla oblongata and descends on the dorsolateral surface of the
spinal cord posterior to the spinal roots. In its downward course to the cauda,
equina the posterior spinal artery receives a succession of small arterial
branches, which enter the spinal canal through the intervertebral foramina.
These vessels and their branches anastomose freely around the posterior roots
and with the corresponding vessels on the opposite side, dipping also into the
substance of the spinal cord, and in the midline form the posterior central
artery.
13.4.5.3 The anterior spinal
artery is formed by the union of two branches from the terminal portion of the
vertebral artery at the level of the foramen magnum. The artery descends as a
single trunk on the anterior aspect of the spinal cord to the conus medullaris,
then continues along the cauda equina and ends as a fine arteriole accompanying
the filum terminale. At successive levels spinal branches entering through the
intervertebral foramina, reinforce it. Along its course, small twigs from this
artery enter the sub- stance of the spinal cord and in the anterior median
fissure these form the anterior central artery.
13.4.5.4 The spinal branches arise
at various levels from the sacral, iliolumbar, intercostal, inferior thyroid
and vertebral arteries, which enter the spinal canal through the intervertebral
foramina. Each spinal branch divides into two rami.
13.4.5.5 A peripheral ramus, which
after entering the spinal canal divides into an ascending and descending
branch. It then anastomoses with the one above and below to form two lateral
chains on the posterior surfaces of the vertebral bodies near the junction of
the pedicles.
13.4.5.6 (2) A central ramus,
which supplies the spinal cord and its membranes by dividing into anterior and
posterior arteries, which anastomose with the anterior and posterior arteries
of the spinal cord.
13.4.6
Venous Drainage of Spinal Cord and Vertebral Column Illustration # 6
Ciba 54
13.4.6.1 Outside and inside the
vertebral canal, running along the entire length, is a series of venous
plexuses which freely join with each other and end in intervertebral veins.
13.4.6.2
Two groups of venous plexuses are found on the outside of the vertebral
canal: (I) the anterior group lies in front of the vertebral bodies. This group
receives some venous tributaries from vertebral bodies and communicates with
the basivertebral and intervertebral veins, and (2) the posterior group, which
forms a network of venous plexuses spreading over the spinous processes,
laminae, facets and adjacent deep musculature. In the cervical region, these
veins communicate with the deep cervical, occipital and cerebral veins.
13.4.6.3 The venous plexuses on
the inside of the vertebral canal lie between the dura and inner vertebral
surfaces. These veins receive tributaries from the adjacent bony structures and
the spinal cord. Although they form a close network, running vertically within
the spinal canal, they may be subdivided as follows.
13.4.6.4 A pair of anterior
internal venous plexuses which lie on either side of the posterior longitudinal
ligament and into which basivertebral veins empty.
13.4.6.5 (2) A single posterior
internal venous plexus which lies anterior to, and on either side of, the vertebral
arches and ligamentum flavum, and anastomose with the posterior external veins.
13.4.6.6 These plexuses form
almost a series of venous rings at the level of each vertebra, found most
strikingly at the foramen magnum. Tunneling the bony structure of each vertebral
body is the basivertebral vein, which has a small valve-like opening as it
joins the anterior internal venous plexus.
13.4.6.7 The intervertebral veins
leave the spinal cord through the intervertebral foramina in company with the
spinal nerves. They also have a valve-like mechanism as they join the
intercostal, lumbar and sacral veins.
13.4.6.8 The veins of the spinal cord are minute and delicate. They emerge from the anterior median fissure as the anterior central vein, and from the posterior sulcus as the posterior central vein. There are also two lateral longitudinal veins on either side of the spinal cord, and they all empty into the intervertebral veins. How- ever, those near the foramen magnum empty into the inferior petrosal sinus or cerebellar veins.
13.5 Exit of Spinal Nerves Ciba 50
13.5.1
As the anterior root of the spinal cord emerges from the anterior and
lateral gray columns, it traverses the surrounding membranes of pia, arachnoid
and dura. The posterior root, which is attached to the posterolateral portion
of the spinal cord, originates from two bundles of fibers in the spinal
ganglion. Both anterior and posterior roots pierce the dura separately as they
make their exit through their respective intervertebral foramina. As a rule,
the posterior root is thicker and larger than the anterior root. They are
enclosed in a common dural sheath just beyond the spinal ganglion, where they
become the spinal nerve and are surrounded by epineurium.
13.5.2
The spinal ganglia, which lie at the outer portion of the intervertebral
foramina, are oval-shaped and vary in size corresponding to their nerve roots.
13.5.3
The Spinal nerves lie horizontally in the cervical region but below these
segments the spinal nerves assume an increasingly oblique and downward
direction as they approach the lumbar region where they are almost vertical,
forming the cauda equina. At the lower thoracic level, there is a
difference of two vertebral segments between the origin of the spinal nerve and
the level of exit. From each sympathetic trunk ganglion, which lies on the
posterolateral surface of the vertebral body, a branch (gray ramus communicans)
joins the adjacent spinal nerve.
13.5.4
Efferent, preganglionic sympathetic fibers (white ramus communicans),
which originate in the lateral column, pass along with the anterior root to the
corresponding sympathetic ganglion or along its trunk to sympathetic plexuses.
13.5.5
Shortly after emerging from the intervertebral foramen each spinal nerve
gives off a meningeal branch, which turns back through the same foramen to
supply the spinal cord membranes, blood vessels, intervertebral ligaments and
joint surfaces.
13.5.6
The spinal nerve then divides into two branches, each with fibers from
both roots: I. Anterior division supplies the anterior and lateral portions of
the trunk and limbs.
13.5.7
In the thoracic region it spans the space between the pleura and
intercostal membrane, runs below the lower rib margin and supplies the
intercostal muscles and adjacent skin.
13.5.8
In the cervical and lumbar regions the anterior divisions form plexuses.
2. Posterior division is directed backward shortly beyond the formation of the
spinal nerve. Its medial branch Supplies the multifides, longissimus,
semispinalis and trapezius, then proceeds along the spinous process, and
supplies the skin. Its lateral branch traverses the longissimus and supplies
the intercostal muscle and adjacent skin.
13.5.9 In the lumbar region the medial branches of the posterior division hug the articular processes of the vertebrae and end in the multifides, and the lateral branches supply the group of sacrospinalis muscles, adjacent fascia and skill.
13.6
Posterior aspect of the cervical segment of the dura mater (right
half) Photo At 69
13.6.1
To demonstrate the deep structures contained within the vertebral
column) the following steps are carried out:
13.6.2
All muscles from the back of neck are removed
13.6.3
The vertebra/laminae) together with the spinous processes) are
removed.
13.6.4
The content of the epidural space (connective tissue and venous
plexus) is removed from the posterior aspect of the dura mater
13.6.5
The intervertebral foramina are enlarged posteriorly
13.6.6
Several vertebrarterial foramina are opened posteriorly.
13.6.7 Following such a dissection the extensions of the dura mater to the spinal ganglia, distribution of spinal nerves into the rami, grouping of the ventral rami, and the relationship of the vertebral artery to the transverse processes and the spinal nerves become 'apparent.
13.7 Spinal Nerves ??
13.7.1 Nerves are such delicate things if you look at them laid bare by the scalpel. A spider web latticework connects our brain to the rest of our body. When the connection is broken, we loose sensation and function. When nerves are compressed or irritated by our own body’s chemicals we ache and moan. They cause the muscles to bunch up tight into a knot, which begs for release.
13.7.2 There are two basic kinds of nerves, sensory and motor. Sensory nerves supply the body sensation and motor nerves innervate the muscles and allow you to move. The structure of these nerves and the way they exit the spinal cord and supply the rest of the body with sensation and movement is significant because they pass through structures, which can irritate and cause pain. In addition, depending on which part of the nerve is irritated may determine where the pain is felt.
13.7.3 The spinal roots exit the spinal cord as either sensory (posterior root) or motor (anterior root). Each dorsal root has a swelling (ganglion) outside the spinal cord. The ganglion contains the cell bodies (neurons) whose dendrites transmit stimulation from the periphery. The axon continues from the ganglion to the spinal cord on its way to the brain. Motor nerves contain axons and sensory nerves consist of dendrites. The spinal nerves combine and exit the spine via the intervertebral foramen. Shortly after exiting the spine these combined sensory and motor nerves form the anterior ramus, posterior ramus) and meningeal branches. The anterior and posterior rami both contain sensory and motor fibers.
13.7.4 The posterior ramus innervates the muscles of the back, branches into the cutaneous nerves that supply the skin of the back, and branch into the dermatomes. The dermatomes and cutaneous nerves overlap each other in supplying the skin with sensitivity, more or less supplying the same areas at each spinal level.
13.7.5 The anterior rami of the spinal nerve innervate the muscles, joints, and skin of the extremities. the anterior ramus also supplies cutaneous sensitivity to the lateral and anterior trunk.
13.7.6 The meningeal branch re-enters the spinal canal through the intervertebral foramen and supplies the vertebrae, vertebral ligaments, blood vessels of the spinal cord, and the meninges.
13.7.8
Spinal nerves are the paths of communication
between the brain, spinal cord tracts and the periphery.
13.7.9
Each pair of spinal nerves is connected to a
segment of the cord by two points of attachment called roots.
13.7.10 The
posterior dorsal (sensory) root contains sensory nerve fibers only and
conducts nerve impulses from the periphery to the spinal cord.
13.7.11 Each
dorsal root also has a swelling the posterior or dorsal (sensory) root
ganglion, which contains the cell bodies of the sensory neurons from the
periphery.
13.7.12 The
other point of attachment of the spinal nerve to the cord is the anterior or
ventral (motor) root. It contains motor neuron axons only and conducts
impulses from the spinal cord to the periphery.
13.7.13 These
two roots motor and sensory form one nerve, which then exits the spine via the
intervertebral foramen.
13.7.14 Shortly
after a spinal nerve leaves its intervertebral foramen, it divides into several
branches. These branches are known as rami (RA-mi).
13.7.15 The
dorsal ramus (Ramus) innervates the deep muscles and skin of the dorsal
surface of the back.
13.7.16 The
ventral ramus of a spinal nerve innervates the muscles and structures of
the extremities and the lateral and ventral trunk.
13.7.17 In
addition to dorsal and ventral rami, spinal nerves also give off a meningeal
branch.
13.7.18
This branch reenters the spinal canal through
the intervertebral foramen and supplies the vertebrae, vertebral ligaments,
blood vessels of the spinal cord, and the meninges.
13.7.19
13.8
Disorders:
Homeostatic Imbalances
13.9
The
Brain and Cranial Nerves
13.10
The
Sensory, Motor and Integrative Systems
13.11
The
Autonomic Nervous System
13.12
The
Special Senses
14 The Brain and Cranial Nerves
14.1 Principal Parts
14.1.1 The brain is mushroom shaped, weighs about 3 lbs., is composed of 1,000,000,000,000 (1 trillion) neurons and as such is one of the largest organs in the body.[6] It is divided into four parts the brain stem, diencephalon, cerebrum, and cerebellum. The brain stem, which is the stock of the mushroom, consists of the medulla oblongata, pons, and midbrain. Superior to brain stem is a diencephalon which contains the thalamus and hypothalamus. The cerebrum accounts for most of the weight of the brain (7/8th) and occupies most of the cranium. Underneath this cerebrum and behind the brainstem is the cerebellum.
14.2 Protection and Coverings
14.2.1 The brain has the same protective coverings as the spinal cord e.g. dura mater, arachnoid, and pia mater. The cranial dura mater consists of two layers. The outer layer is thicker and adheres tightly to the cranial bones. This outer layer is not found covering the spinal cord. The dura maters second meningeal layer is contiguous with the meningeal layer of the spinal cord.
14.3 Cerebrospinal fluid and ventricles
14.3.1 There are four ventricles, two of which are called the lateral ventricles. The lateral ventricles are on either side of the right and left halves of the brain. These two ventricles are connected on both sides by the interventricular foramen to the third ventricle. The third ventricle is a vertical slit between both halves of the thalamus an between the two lateral ventricles. The fourth ventricle is connected by the cerebral aqueduct to the third ventricle. The fourth ventricle is connected to the sub arachnoid space via the two lateral apertures and to the central canal via median aperture. Contained in the ventricles, the central canal, and sub-arachnoid space is 3-5 oz. of cerebrospinal fluid. The cerebrospinal fluid is composed of proteins, glucose, urea, lymphocytes, and salts. Cerebrospinal fluid is exchanged between capillaries and ependymal cells, which form plexuses along the roof of all of the ventricles. The lining in these areas of the ventricles is composed of ependymal cells (specialized neuroglial cells that form the lining of all of the ventricles) and capillaries, which form these plexuses called choroid plexuses. These choroid plexuses allow exchange of proteins, glucose, urea, lymphocytes, and salts between the bloodstream and ventricles. The byproducts of the metabolism of brain cells are filtered into the bloodstream for removal and nutrients necessary for healthy brain cells are exchanged from the bloodstream to the ventricles. the quarry plexuses form and effective Lund cerebrospinal fluid barrier that prevents coccyx up to 30 from entering the ventricles but. Can nutrients. The blood brain barrier or blood cerebrospinal fluid barrier he's a protective mechanism in each ventricles to prevent substances from harming midbrain. One possible schematic for the circulation of cerebrospinal fluid (CSF) in the brain is as follows; CSF is produced by the choroid plexuses from the lateral ventricles, which then circulates through the interventricular foramen to the third ventricle. In the 3rd ventricle, more CSF is added by the choroid plexuses flowing to the 4th ventricles via the cerebral Aqueduct. At the 4th ventricles more C S F is added from the choroid plexuses and then continues on to the sub arachnoid space via the lateral aperture where it flows around the back of the brain and posterior spinal cord. Cerebrospinal fluid also can flow through the median aperture both to the sub arachnoid space around the anterior portion of brain and the sub arachnoid space around the anterior portion of the spinal cord. From the median aperture, CSF can flow down the central canal of the spinal cord. Cerebrospinal fluid is gradually absorbed into the veins after circulating through the ventricles. Additional cerebrospinal fluid can also be formed by ependymal (neuroglial) cells in a central canal the spinal cord. This flows up the central canal to be added to the full in the 4th ventricles. Cerebrospinal fluid is absorbed into the veins via the arachnoid using finger like projections called arachnoid villi. These finger like projections push through dural venous sinuses such as the superior sagittal sinus. CSF is absorbed as fast as it is produced.
14.3.2 When CSF is prevented from flowing because of an obstruction in the ventricles for in the sub arachnoid space it is called hydrocephalus. Internal hydrocephalus is caused from a blockage in the ventricles and external hydrocephalus is caused by blockage of the arachnoid space.
14.3.3 One principal function of the cerebrospinal fluid are to serve as a shock-absorbing medium to protect the brain & spinal cord from injury, which would otherwise cause them to crash against the bony walls of the cranium and vertebral canal. The second principal function of the cerebrospinal fluid is to deliver nutrition to the brain cells and remove waste and toxic substances.
14.4
Blood Circulation
14.4.1 The cerebrospinal circle (Circle of Willis) supplies the brain with oxygen and nutrients. The blood vessels that pass along the outer surface of the brain and penetrate inward are surrounded by pia mater. The space between the pia mater and the penetrating blood vessel is called perivascular space. The brain uses 20% of the oxygen supply utilized by the entire body yet represents only 2% of its weight. The brain is very active metabolically and can only tolerate short periods without the necessary supply of oxygen. Four minutes of oxygen, deprivation may result in permanently injured brain cells. Additionally the lysosomes in brain cells release enzymes, which bring about the self-destruction of the cell. Babies deprived of oxygen while going thru the birth canal can suffer mental retardation, epilepsy, and or paralysis. Because glucose is the primary fuel of the brain cells deprivation can result in mental confusion, dizziness, convulsions and unconsciousness. -
14.4.2 When the brain is the deprived of oxygen, there is an increase in carbon dioxide (CO2 O2). Both deprivation of oxygen and increase in carbon dioxide can create can increase asset dilation of the blood vessels to the brain. Carbon dioxide does this by combining with water to form carbonic acid (H2CO3). Carbonic acid breaks down further into hydrogen ions (H+) bicarbonate ions (HCO3). The hydrogen ions (H+) increased vasodilatation of the cerebral vessels.
14.4.3 The blood brain barrier (BBB) allows certain substances such as glucose and oxygen to rapidly pass from the capillaries into the brain cells. Capillaries of the brain are constructed differently than other capillaries in the body. Their cells and more densely packed and large numbers of astrocytes (a type of neuroglial cell) are also present. These astrocytes produce substance, which influence the capillaries and increase or decrease their permeability.
14.5
Brain Stem
14.5.1
Medulla Oblongata (Medulla)
14.5.1.1 The medulla is about an inch long and lies just above the foramen magnum. As such, it is a continuation the spinal cord and serves as a relay station for all nerve tracks passing to and from the spinal cord. Some of these ascending and he descending nerve tracks cross to the opposite side as they pass through medulla. The largest of these nerve tracks pass on the anterior side of the medulla and form pyramids that cross to the opposite side and are called the decussation of the pyramids. This is why motor movements of one side of the body are controlled by the opposite side of the brain. There is no known adaptive value for these phenomena. There is a region of gray matter in the medulla called the reticular formation. The reticular formation is also located in the spinal cord, Pons, midbrain and diencephalon. The reticular formation connects to many parts of the brain and is involved in arousal from sleep. The medulla also contains some of the cranium nerves.
14.5.1.2 The vestibulocochlear (VIII) nerve is concerned with hearing and equilibrium; the glossopharyngeal (IX) nerve is concerned with swallowing, salvation, and taste; the vagus (X) relays nerve impulses to from many of the thoracic and abdominal viscera; the cranial portion of the accessory (XI) nerve originates in the 5 cervical vertebrae of the spinal cord. The accessory (XI) nerve conveys nerve impulses related to head and shoulder movements. The glossopharyngeal (IX) convey nerve impulses that involve tongue movements.
14.5.1.3 The inferior accessory olivary nucleus projects nerve fibers to the cerebellum. The inferior olivary nucleus is concerned with efficiency of voluntary movements especially precision ones. The accessory olivary nucleus is concerned with equilibrium, postural changes, and locomotion. The vestibular nuclear complex maintains a sense of equilibrium.
14.5.1.4 There are three reflex centers in the medulla; the cardiac center, which regulates the rate of heart, beat the force of contraction; the medullary rhythmicity area, which adjusts the basic rhythm of breathing; and the vasoconstrictor center, which regulates the diameter of blood vessels. The medulla also has centers, which control following, vomiting, coughing, sneezing, and hiccuping.
14.5.2
Pons
14.5.2.1 The Pons (Bridge) is about an inch long and as the name implies connects the spinal cord with the brain and parts of the brain with each other. It lies directly above the medulla and is anterior to the cerebellum. The connections it makes in the brain are in two principal directions e.g. the transverse fibers connect with the cerebellum and the longitudinal fibers of the Pons belong to the motor and sensory tracks that connect the spinal cord for medulla with the other parts of the brain stem.
14.5.2.2 The following cranial nerves have neurons, which originate in the Pons. The trigeminal (V) nerve controls impulses for chewing and for sensations of the head and face. The abducens (VI) nerve regulates certain, eyeball movements. The facial (VII) nerve, which conducts impulses, related to taste, salvation, and facial expression. The vestibulocochlear (VIII) nerve (vestibular branches), is concerned with equilibrium.
14.5.2.3 The pneumotaxic area and apneustic area together with the medullary rhythmicity area in the medulla control respiration (breathing movements).
14.5.2.4 Oculomotor (III) trochlear (IV) lemniscus mesencephalon that the tectum corpora quadrigemina colliculi substanti nigra
14.5.3 Midbrain
14.5.3.1 The midbrain extends from the pons to the lower portion of the diencephalon.
14.6
Diencephalon
14.6.1 The diencephalon consists principally of this thalamus and hypothalamus.
14.6.2
Thalamus
14.6.2.1 This thalamus is an oval structure above the midbrain that measures about 1 in. in length and constitutes 4/5 of the diencephalon.
14.6.3
Test your Knowledge
Click
the link below to take the section test.
14.6.4
Hypothalamus
14.7
Cerebrum
14.7.1
Lobes
14.7.2
White
Matter
14.7.3
Basal
Ganglia (Cerebral Nuclei)
14.7.4
Limbic
System
14.7.5
Functional
Areas of Cerebral Cortex
14.7.6
Electroencephalogram
(EEG)
14.8
Brain
Lateralization (Split-Brain Concept)
14.9
Cerebellum
14.9.1
Structure
14.9.2
Functions
14.10
Neurotransmitters
in the Brain
14.10.1
Neurotransmission
Merck
14.10.1.1 General
14.10.1.1.1
A
nerve cell (neuron) has two major functions: propagation of an action potential
(nerve impulse, signal) along its axon and transmission of this signal from one
neuron to another neuron or to an effector cell to elicit a response. Effector
cells include skeletal and cardiac muscles and exocrine and endocrine cells
regulated by the nervous system. Impulse conduction along an axon is
electrical, caused by the exchanges of Na+ and K+ ions
across the neuronal membrane. In contrast, impulse transmission from one neuron
to another neuron or to a non-neuronal effector cell depends on the action of
specific neurotransmitters on specific receptors.
14.10.1.1.2
A
particular neuron generates an identical action potential after each stimulus
and conducts it at a fixed velocity along the axon. Velocity depends on axonal
diameter and degree of myelination. For myelinated fibers, velocity (m/sec) is
approximately 3.7 times the diameter (µ); eg, for a large (20-µ) myelinated
fiber, velocity is nearly 75 m/sec. For unmyelinated fibers with diameters of 1
to 4 µ, velocity is 1 to 4 m/sec.
14.10.1.1.3
A
neuron simultaneously receives many stimuli--positive and negative--from other
neurons and integrates them into various patterns of firing. The nerve impulses
travel down axons to the next synapse. Once axonal propagation begins, drugs or
toxins can modify the amount of neurotransmitter released from the terminal
axon. For example, botulinum toxin blocks release of acetylcholine. Other
chemical factors can also influence the effect of transmission by modifying a
receptor. In myasthenia gravis, antibodies block the nicotinic acetylcholine
receptor.
14.10.1.1.4
Synapses
occur between neuron and neuron and, in the periphery, between neuron and
effector (eg, a muscle); in the CNS, more complex arrangements exist.
Functional contact between two neurons may occur between axon and cell body,
axon and dendrite (the receiving area of a neuron), cell body and cell body, or
dendrite and dendrite. Neurotransmission can increase or decrease to generate a
physiologic function or to respond to changing physiologic needs. Many
neurologic and psychiatric diseases are caused by pathologic overactivity or underactivity
of neurotransmission. Many drugs can modify neurotransmission; some (eg,
hallucinogens) cause adverse effects, and some (eg, antipsychotic drugs)
correct pathologic conditions.
14.10.1.1.5 Development and maintenance of cells in the nervous system depend on many specific proteins, such as nerve growth factor, brain-derived neurotrophic factor, and neurotrophin-3.
14.10.1.2 Basic Principles of Neurotransmission
14.10.1.2.1
The
nerve cell body produces enzymes that are involved in the synthesis of most
neurotransmitters. The enzymes act on precursor molecules taken up by the
neuron to form the neurotransmitter. The neurotransmitter is stored at the
nerve terminal in vesicles (see Fig. 166-1). The amount in one vesicle (usually several thousand
molecules) is a quantum. Some neurotransmitter molecules are constantly
extruded from the terminal, but the amount is insufficient to elicit a
significant physiologic response. An action potential arriving at the terminal can
activate calcium currents and precipitate the simultaneous release of
neurotransmitter molecules from many vesicles by fusing the membrane of the
vesicles to that of the nerve terminal. An opening forms through which the
molecules are expelled into the synaptic cleft by exocytosis.
14.10.1.2.2
The
quantity of neurotransmitters in the terminals is kept relatively constant and
independent of nerve activity by tight regulation of neurotransmitter
formation. Regulation varies among neurons and is achieved by modifying precursor
uptake or the activity of enzymes involved in neurotransmitter synthesis or
destruction. Stimulation or blockade of postsynaptic receptors can decrease or
increase presynaptic neurotransmitter synthesis.
14.10.1.2.3
Neurotransmitters
diffuse across the synaptic cleft, bind briefly to receptors, and activate
them, causing physiologic responses. Depending on the receptor, the response
may be excitatory (ie, initiating a new action potential) or inhibitory (ie,
inhibiting the development of a new action potential).
14.10.1.2.4
The
neurotransmitter-receptor interaction must be terminated quickly so that the
same receptors can be rapidly activated repeatedly. The neurotransmitter is
quickly pumped back into the presynaptic nerve terminals by active processes
(reuptake), is destroyed by enzymes near the receptors, or diffuses into the
surrounding area and is destroyed.
14.10.1.2.5 Abnormalities of neurotransmitter synthesis, storage, release, or degradation or changes in the number and affinity of receptors can affect neurotransmission and cause clinical disorders (see Table 166-1).
14.10.1.3 Major Neurotransmitters
14.10.1.3.1
A
neurotransmitter is a chemical that is selectively released from a nerve
terminal by an action potential, interacts with a specific receptor on an
adjacent structure, and, if received in adequate amounts, elicits a specific
physiologic response. To be a neurotransmitter, a chemical must be present in
the nerve terminal, be released from the nerve terminal by an action potential,
and, when applied experimentally to the receptor, always produce the identical
effect. Many chemicals act as neurotransmitters. There are at least 18 major
neurotransmitters; several occur in slightly different forms.
14.10.1.3.2 The amino acids glutamate and aspartate are the major excitatory neurotransmitters in the CNS. They occur in the cortex, cerebellum, and spinal cord.
14.10.1.3.3
Aminobutyric
acid (GABA)
14.10.1.3.3.1 major inhibitory neurotransmitter in the brain. GABA is derived from glutamic acid, which is decarboxylated by glutamate decarboxylase. After interaction with its receptors, GABA is actively pumped back into the nerve terminals and metabolized. Glycine, which resembles GABA in its action, occurs principally in the interneurons of the spinal cord. Glycine is probably metabolized from serine.
14.10.1.3.4 Serotonin (5-hydroxytryptamine, or 5-HT)
14.10.1.3.4.1 generated by the raphe nucleus and midline neurons of the pons and upper brain stem. Tryptophan is hydroxylated by tryptophan hydroxylase to 5-hydroxytryptophan, then decarboxylated to serotonin. Serotonin levels are controlled by the uptake of tryptophan and intraneuronal monoamine oxidase.
14.10.1.3.5
Acetylcholine
14.10.1.3.5.1 major neurotransmitter of the bulbospinal motor neurons, autonomic preganglionic fibers, postganglionic cholinergic (parasympathetic) fibers, and many neurons in the CNS (eg, basal ganglia, motor cortex). It is synthesized from choline and mitochondrially derived acetyl coenzyme A by choline acetyltransferase. When released, acetylcholine stimulates specific cholinergic receptors, an interaction rapidly terminated by local hydrolysis of acetylcholine to choline and acetate by acetylcholinesterase. Acetylcholine levels are regulated by choline acetyltransferase and by choline uptake.
14.10.1.3.6
Dopamine
14.10.1.3.6.1 neurotransmitter of some peripheral nerve fibers and many central neurons (eg, in the substantia nigra, midbrain, ventral tegmental area, and hypothalamus). The amino acid tyrosine is taken up by dopaminergic neurons and converted by tyrosine hydroxylase to 3,4-dihydroxyphenylalanine (dopa), which is decarboxylated by aromatic-L-amino-acid decarboxylase to dopamine. After release, dopamine interacts with dopaminergic receptors, and its residue is actively pumped back (reuptake) into the prejunctional neurons. Tyrosine hydroxylase and monoamine oxidase regulate dopamine levels in nerve terminals.
14.10.1.3.7
Norepinephrine
14.10.1.3.7.1
neurotransmitter
of most postganglionic sympathetic fibers and many central neurons (eg, in the
locus caeruleus and hypothalamus). The precursor tyrosine is converted to
dopamine, which is hydroxylated by dopamine  -hydroxylase
to norepinephrine. When released, norepinephrine interacts with adrenergic
receptors, a process terminated by its reuptake into the prejunctional neurons
with subsequent degradation by monoamine oxidase and by the action of catechol O-methyltransferase (COMT), which is
located mainly extraneuronally. Tyrosine hydroxylase and monoamine oxidase
regulate intraneuronal norepinephrine levels.
-hydroxylase
to norepinephrine. When released, norepinephrine interacts with adrenergic
receptors, a process terminated by its reuptake into the prejunctional neurons
with subsequent degradation by monoamine oxidase and by the action of catechol O-methyltransferase (COMT), which is
located mainly extraneuronally. Tyrosine hydroxylase and monoamine oxidase
regulate intraneuronal norepinephrine levels.
14.10.1.3.8
Endorphin
14.10.1.3.8.1
and other
endorphins are polypeptides that activate many central neurons (eg, in the
hypothalamus, amygdala, thalamus, and locus caeruleus). The cell body contains
a large polypeptide called pro-opiomelanocortin, the precursor of several
neuropeptides (eg,  -,
-,
and
-,
-,
and  -endorphins).
This polypeptide is transported down the axon and is cleaved into specific
fragments; one is -endorphin,
which contains 31 amino acids. After release and interaction with opioid
receptors, -endorphin
-endorphins).
This polypeptide is transported down the axon and is cleaved into specific
fragments; one is -endorphin,
which contains 31 amino acids. After release and interaction with opioid
receptors, -endorphin
14.10.1.3.9
Met-enkephalin and leu-enkephalin
14.10.1.3.9.1 small peptides present in many central neurons (eg, in the globus pallidus, thalamus, caudate, and central gray matter). Their precursor, proenkephalin, is formed in the cell body, then split by specific peptidases into smaller peptides. The resulting fragments include two enkephalins, each composed of five amino acids and containing either methionine or leucine terminals. After release and interaction with peptidergic receptors, the enkephalins are hydrolyzed into smaller, inactive peptides and amino acids, as are dynorphins and substance P.
14.10.1.3.10
Dynorphins
14.10.1.3.10.1 a group of seven peptides with similar amino acid sequences. They geographically coexist with enkephalins. Substance P, a peptide, occurs in central neurons (habenula, substantia nigra, basal ganglia, medulla, and hypothalamus) and in high concentration in the dorsal root ganglia. It is released by intense afferent painful stimuli.
14.10.1.3.10.1.1 Neurotransmitters whose roles in neurotransmission have been less firmly established include histamine, vasopressin, vasoactive intestinal peptide, carnosine, bradykinin, cholecystokinin, bombesin, somatostatin, corticotropin releasing factor, neurotensin, and possibly adenosine.
14.10.1.4 Major Receptors
14.10.1.4.1
Neurotransmitter
receptors are protein complexes that span the cell membrane. Receptors coupled
to a second messenger are usually monomeric and consist of three parts: the
extracellular part, where glycosylation occurs; the transmembrane part, which
forms a pocket where the neurotransmitter is presumed to act; and the
intracytoplasmic part, where G-protein binding or regulation by phosphorylation
of the receptor occurs. Ion channel receptors are multimeric. In some cases,
activation of the receptor results in modification of channel ion permeability.
In others, activation of a second messenger results in changes of channel
conductance.
14.10.1.4.1.1
Receptors
that are continuously stimulated by neurotransmitters or drugs (agonists)
become hyposensitive (down-regulated); those that are not stimulated by their
neurotransmitter or are blocked chronically by drugs (antagonists) become
hypersensitive (up-regulated). Up- or down-regulation of receptors strongly
influences the development of tolerance and physical dependence. Withdrawal is
usually a rebound phenomenon due to altered receptor affinity or density. These
concepts are particularly important in organ or tissue transplants, in which
the receptors are deprived of the physiologic neurotransmitter by denervation.
14.10.1.4.1.1.1 Most neurotransmitters interact primarily with postsynaptic receptors, but some receptors are located on presynaptic neurons, providing fine control of neurotransmitter release.
14.10.1.4.2
Cholinergic receptors
classified as nicotinic N1 (in the adrenal medulla and autonomic ganglia) or N2 (in skeletal muscle) or muscarinic M1 (in the autonomic nervous system, striatum, cortex, and hippocampus) or M2 (in the autonomic nervous system, heart, intestinal smooth muscle, hindbrain, and cerebellum).
14.10.1.4.3
Adrenergic receptors
classified
as ![]() 1
(postsynaptic in the sympathetic system),
1
(postsynaptic in the sympathetic system), ![]() 2
(presynaptic in the sympathetic system and postsynaptic in the brain),
2
(presynaptic in the sympathetic system and postsynaptic in the brain), ![]() 1
(in the heart), or
1
(in the heart), or ![]() 2
(in other sympathetically innervated structures). Dopaminergic
receptors are classified as D1, D2, D3,
D4, and D5. D3 and D4 play a role
in thought control (limit negative symptoms of schizophrenic processes),
whereas D2 receptor activation controls the extrapyramidal system.
2
(in other sympathetically innervated structures). Dopaminergic
receptors are classified as D1, D2, D3,
D4, and D5. D3 and D4 play a role
in thought control (limit negative symptoms of schizophrenic processes),
whereas D2 receptor activation controls the extrapyramidal system.
14.10.1.4.4
GABA receptors
classified as GABAA (activating chloride channels) and GABAB (potentiating cAMP formation). The GABAA receptor consists of several distinct polypeptides and is the site of action for several neuroactive drugs, including benzodiazepines, newer anticonvulsants (eg, lamotrigine), barbiturates, picrotoxin, and muscimol.
14.10.1.4.5
Serotoninergic (5-HT) receptors
(with at least 15 subtypes) are classified as 5-HT1 (with four subtypes), 5-HT2, and 5-HT3. 5-HT1A receptors, which occur presynaptically in the raphe nucleus (inhibiting presynaptic uptake of 5-HT) and postsynaptically in the hippocampus, modulate adenylate cyclase. 5-HT2 receptors, located in the fourth layer of the cortex, are involved in phosphoinositide hydrolysis (see Table 166-2). 5-HT3 receptors occur presynaptically in the nucleus tractus solitarius.
14.10.1.4.6
Glutamate receptors
classified as ionotrophic NMDA (N-methyl-D-aspartate) receptors, which bind NMDA, glycine, zinc, Mg++, and phencyclidine (PCP, also known as angel dust) and affect the influx of Na+, K+, Ca++, and non-NMDA receptors, which bind quisqualate and kainate. Non-NMDA channels are permeable to Na+ and K+ but not to Ca++. These excitatory receptors mediate important toxic effects by increasing calcium, free radicals, and proteinase. In neurons, synthesis of nitric oxide (NO) involving NO synthase increases in response to glutamate.
14.10.1.4.7
Endorphin-enkephalin (opioid) receptors
classified
as µ1 and µ2 (affecting sensorimotor integration and
analgesia), ![]() 1
and
1
and ![]() 2
(affecting motor integration, cognitive function, and analgesia), and
2
(affecting motor integration, cognitive function, and analgesia), and ![]() 1,
1,
![]() 2,
and
2,
and ![]() 3
(affecting water balance regulation, analgesia, and food intake). Sigma
receptors, currently classified as nonopioid and mostly localized in the
hippocampus, bind PCP.
3
(affecting water balance regulation, analgesia, and food intake). Sigma
receptors, currently classified as nonopioid and mostly localized in the
hippocampus, bind PCP.
14.10.1.5 Neurotransmitter Transport
14.10.1.5.1 Two types of neurotransmitter transporters are essential for neurotransmission. The uptake carrier, located on presynaptic neurons and plasma cells, pumps neurotransmitters from the extracellular space into the cell. It replenishes the neurotransmitter supply, helps terminate the action of the neurotransmitter, and, in the case of glutamate, keeps the amount of neurotransmitter below toxic levels. The energy required by these pumps is provided by ATP. The other type of transporter, located on the membrane of the vesicle, concentrates the neurotransmitter into the vesicles for further exocytosis. These transporters are powered by cytoplasmic pH and the voltage gradient across the vesicular membrane. During anoxia or ischemia, the transmembrane ionic gradient changes, and glutamate is transported from the vesicles into the cytoplasm, increasing the intracellular concentration of glutamate to potentially lethal levels.
14.10.1.5.2
Second messenger systems
14.10.1.5.2.1 consist of regulatory G proteins and catalytic proteins (eg, adenylate cyclase, phospholipase C) that link receptors and effectors. A second messenger can be the trigger of a chain reaction or the target of a regulatory pathway (eg, calcium--see Table 166-2).
14.11
Cranial
Nerves
14.12
Aging
and the Nervous System
14.13
Developmental
Anatomy of the Nervous System
15 Sensory, Motor, and Integrative Systems
16 Autonomic Nervous System
17 Special Senses
18 Endocrine System*
18.1
Endocrine
Glands
18.2
Chemistry
of Hormones
18.3
Physiology
of Hormonal Action
18.3.1
Overview
18.3.2
Receptors
and Hormone Specificity
18.3.3
Interaction
with Plasma Membrane Receptors
18.3.4
Interaction
with Intracellular Receptors
18.3.5
Hormonal
Interactions
18.3.6
Prostaglandins
(PGs) and Hormones
18.4
Control
of Hormonal Secretions: Feedback Control
18.5
Pituitary
(Hypophysis)
18.5.1
Adenohypophysis
18.5.1.1
Human
Growth Hormone (hGH)
18.5.1.2
Thyroid
Stimulating Hormone (TSH)
18.5.1.3
Adrenocorticotropic
Hormone (ACTH)
18.5.1.4
Follicle
Stimulating Hormone (PSH)
18.5.1.5
Luteinizing
Hormone (LH)
18.5.1.6
Prolactin
(PRL)
18.5.1.7
Melanocyte
Stimulating Hormone (MSH)
18.5.2
Neurohypophysis
18.5.2.1
Oxytocin
(OT)
18.5.2.2
Antidiuretic
Hormone (ADH)
18.6
Thyroid
18.6.1
Formation,
Storage, and Release of Thyroid Hormones
18.6.2
Function
and Control of Thyroid Hormones
18.6.3
Calcitonin
(CT)
18.7
Parathyroids
18.7.1
Parathyroid
Hormone (PTH)
18.8
Adrenals
(Suprarenals)
18.8.1
Adrenal
Cortex
18.8.1.1
Mineralocorticoids
18.8.1.2
Glucocorticoids
18.8.1.3
Gonadocorticoids
18.8.2
Adrenal
Medulla
18.8.2.1
Epinephrine
and Horepinephrine (NE)
18.9
Pancreas
18.9.1
Glucagon
18.9.2
Insulin
18.10
Ovaries
and Testes
18.11
Pineal
(Epiphusis Cerebrl)
18.12
Thymus
18.13
Aging
and the Endocrine System
18.14
Developmental
Anatomy of the Endocrine System
18.15
Other
Endocrine Tissues
18.16
Growth
Factors
18.17
Stress
and the General Adaptation Syndrome
18.17.1
Stressors
18.17.2
Alarm
Reaction
18.17.3
Resistance
Reaction
18.17.4
Exhaustion
18.17.5
Stress
and Disease
19 Cardiovascular System-Blood*
19.1
Blood
19.1.1
Physical
Characteristics of Blood
19.1.2
Functions
of Blood
19.1.3
Components
of Blood
19.1.3.1
Erthrocytes
19.1.3.2
Leucocytes
19.1.3.3
Thrombocytes
19.1.3.4
Plasma
19.1.4
Physiology
of Hemostosis
19.1.5
Grouping
(Typing) of Blood
19.1.6
Interstitial
Fluid and Lymph
19.1.7
Disorders:
Homeostatic Imbalances
20 Cardiovascular System-Heart
20.1
Location
of Heart
20.2
Pericardium
20.3
Wall;
Chambers; Vessels; and Valves
20.4
Blood
Supply
20.5
Conduction
System
20.6
Electrocardiogram
(ECG)
20.7
Blood
Flow through the Heart
20.8
Physiology
of Cardiac Cycle
20.9
Physiology
of Cardiac Output (CO)
20.10
Artificial
Heart
20.11
Diagnosing
Heart Disorders
20.12
Heart-Lung
Machine
20.13
Risk
Factors in Heart Disease
20.14
Developmental
Anatomy of the Heart
20.15
Disorders:
Homeostatic Imbalances
21 Cardiovascular System-Vessels and Routes
21.1
Arteries
21.2
Arterioles
21.3
Capillaries
21.4
Venules
21.5
Veins
21.6
Blood
Reservoirs
21.7
Physiology
of Circulation
21.8
Shock
and Homeostasis
21.9
Checking
Circulation
21.9.1
Pulse
21.9.2
Measurement
of Blood Pressure
21.10
Circulatory
Routes
21.10.1
Systemic
Circulation
21.10.1.1
Arteries
of Pelvis and Lower Extremities
21.10.1.1.1
External
Iliacs
21.10.1.1.1.1 External Iliac arteries diverge through greater (false) pelvis, enter thighs, and here become right and left femoral arteries.
21.10.1.1.1.2 Both femorals send branches back up to genitals and wall of abdomen.
21.10.1.1.1.3 Other branches run to muscles of thigh.
21.10.1.1.1.4 Femoral continues down medial and posterior side of thigh at back of knee joint, where it becomes the popliteal artery.
21.10.1.1.1.5 Between knee and ankle, popliteal runs down back of leg and is called posterior tibial artery.
21.10.1.1.1.6 Below knee, peroneal artery branches off posterior tibial to supply structures on medial side of fibula and calcaneus.
21.10.1.1.1.7 In calf, anterior tibial artery branches off popliteal and runs along front of leg.
21.10.1.1.1.8 At ankle, it becomes dorsalis pedis artery.
21.10.1.1.1.9 At ankle, posterior tibial divides into medial and lateral plantar arteries.
21.10.1.2
Veins
of Lower Extremities
21.10.1.2.1
Blood from
each lower extremity is returned by superficial set and deep set of veins.
21.10.1.2.2
Superficial
Veins
21.10.1.2.2.1 Great Saphenous
21.10.1.2.2.1.1 This is the longest vein in the body and begins at the medial end of the dorsal venous arch of foot.
21.10.1.2.2.1.2 It passes in front of medial Malleolus and then upward along medial aspect of leg and thigh.
21.10.1.2.2.1.3 It receives tributaries from superficial tissues and connects with deep veins as well.
21.10.1.2.2.1.4 It empties into femoral vein in groin.
21.10.1.2.2.2 Small Saphenous
21.10.1.2.2.2.1 This vein begins at lateral end of dorsal venous arch of foot.
21.10.1.2.2.2.2 It passes behind lateral Malleolus and ascends under skin of back of leg.
21.10.1.2.2.2.3 It receives blood from foot and posterior portion of leg.
21.10.1.2.2.2.4 It empties into popliteal vein behind knee.
21.10.1.2.3
Deep Veins
21.10.1.2.3.1 Posterior Tibial
21.10.1.2.3.1.1 Union of medial and lateral plantar veins behind medial Malleolus forms this vein.
21.10.1.2.3.1.2 It ascends deep in muscle at back of leg, receives blood from peroneal vein, and unites with anterior tibial vein just below knee.
21.10.1.2.3.2 Anterior Tibial
21.10.1.2.3.2.1 This vein is the upward continuation of dorsalis pedis veins in foot.
21.10.1.2.3.2.2 It runs between tibia and fibula and unites with posterior tibial to form popliteal vein.
21.10.1.2.3.3 Popliteal
21.10.1.2.3.3.1 This vein is just behind knee, receives blood from anterior and posterior tibials and small Saphenous vein.
21.10.1.2.3.4 Femoral
21.10.1.2.3.4.1 This vein is the upward continuation of popliteal just above knee.
21.10.1.2.3.4.2 Femorals run up posterior of thighs and drain deep structures of thighs.
21.10.1.2.3.4.3 After receiving great Saphenous veins in groin, they continue as right and left external iliac veins